

Endogenous serotonin acts on 5-HT2C-like receptors in key vocal areas of the brain stem to initiate vocalizations in Xenopus laevis
- PMID: 19955293
- PMCID: PMC2822697
- DOI: 10.1152/jn.00827.2009
Endogenous serotonin acts on 5-HT2C-like receptors in key vocal areas of the brain stem to initiate vocalizations in Xenopus laevis
Abstract
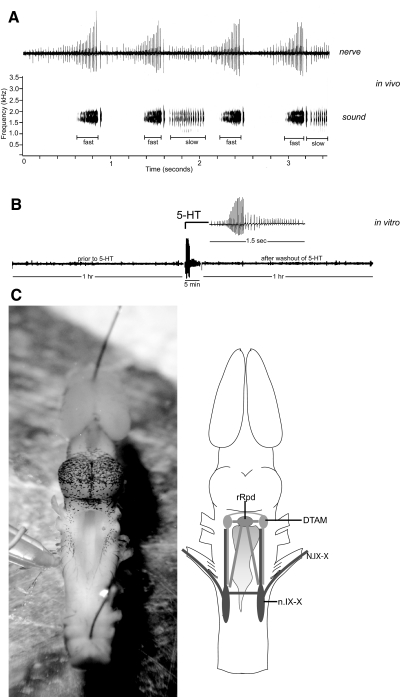
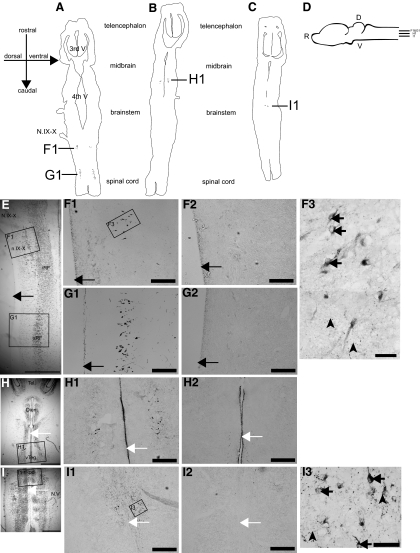
Serotonin initiates various rhythmic behaviors in vertebrates. Previously we have shown that serotonergic neurons innervate the central vocal pathway in the African clawed frog (Xenopus laevis). We also discovered that exogenous serotonin applied to isolated brains in vitro activates fictive vocalizations by activating 5-HT(2C)-like receptors. In this study, we examined the location of 5-HT(2C)-like receptors and determined whether endogenously released serotonin also initiates vocalizations by activating 5-HT(2C)-like receptors in male Xenopus brains. To this end, we first identified the specific location of 5-HT(2C)-like receptors using immunohistochemistry. We next examined which of the populations of neurons that express 5-HT(2C)-like receptors are functionally relevant for initiating fictive vocalizations by applying a 5-HT(2C) receptor agonist to brains transected at various levels. Of four populations of immunopositive neurons, we showed that 5-HT(2C)-like receptors located in two areas of the brain stem vocal circuit, the raphe nucleus and motor nucleus IX-X, initiate fictive vocalizations. We next showed that endogenous serotonin can also activate fictive vocalizations by increasing the extracellular concentration of endogenous serotonin using a selective serotonin reuptake inhibitor (SSRI). The SSRI-induced vocal initiation is also mediated by activation of 5-HT(2C)-like receptors because blockade of these receptors prevents fictive vocalization. The results suggest that in vivo release of serotonin initiates male vocalizations by activating 5-HT(2C)-like receptors in the brain stem vocal nuclei.
Figures




Similar articles
-
Generation, Coordination, and Evolution of Neural Circuits for Vocal Communication.J Neurosci. 2020 Jan 2;40(1):22-36. doi: 10.1523/JNEUROSCI.0736-19.2019. J Neurosci. 2020. PMID: 31896561 Free PMC article. Review.
-
5-HT2C-like receptors in the brain of Xenopus laevis initiate sex-typical fictive vocalizations.J Neurophysiol. 2009 Aug;102(2):752-65. doi: 10.1152/jn.90469.2008. Epub 2009 May 27. J Neurophysiol. 2009. PMID: 19474172 Free PMC article.
-
Xenopus vocalizations are controlled by a sexually differentiated hindbrain central pattern generator.J Neurosci. 2007 Feb 7;27(6):1485-97. doi: 10.1523/JNEUROSCI.4720-06.2007. J Neurosci. 2007. PMID: 17287524 Free PMC article.
-
Rhythm generation, coordination, and initiation in the vocal pathways of male African clawed frogs.J Neurophysiol. 2017 Jan 1;117(1):178-194. doi: 10.1152/jn.00628.2016. Epub 2016 Oct 19. J Neurophysiol. 2017. PMID: 27760822 Free PMC article.
-
Studies on the neuroendocrine role of serotonin.Dan Med Bull. 2007 Nov;54(4):266-88. Dan Med Bull. 2007. PMID: 18208678 Review.
Cited by
-
The Xenopus amygdala mediates socially appropriate vocal communication signals.J Neurosci. 2013 Sep 4;33(36):14534-48. doi: 10.1523/JNEUROSCI.1190-13.2013. J Neurosci. 2013. PMID: 24005304 Free PMC article.
-
Generation, Coordination, and Evolution of Neural Circuits for Vocal Communication.J Neurosci. 2020 Jan 2;40(1):22-36. doi: 10.1523/JNEUROSCI.0736-19.2019. J Neurosci. 2020. PMID: 31896561 Free PMC article. Review.
-
Vocal pathway degradation in gonadectomized Xenopus laevis adults.J Neurophysiol. 2011 Feb;105(2):601-14. doi: 10.1152/jn.00883.2010. Epub 2010 Dec 8. J Neurophysiol. 2011. PMID: 21148092 Free PMC article.
-
Inhibitory and modulatory inputs to the vocal central pattern generator of a teleost fish.J Comp Neurol. 2018 Jun 1;526(8):1368-1388. doi: 10.1002/cne.24411. Epub 2018 Feb 28. J Comp Neurol. 2018. PMID: 29424431 Free PMC article.
-
NMDAR-dependent control of call duration in Xenopus laevis.J Neurophysiol. 2010 Jun;103(6):3501-15. doi: 10.1152/jn.00155.2010. Epub 2010 Apr 14. J Neurophysiol. 2010. PMID: 20393064 Free PMC article.
References
-
- Bem T, Orsal D, Cabalquen JM. Fictive locomotion in the thalamic rat. Exp Brain Res 97: 301–304, 1993 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources