

Common threads in persistent viral infections
- PMID: 19955304
- PMCID: PMC2863747
- DOI: 10.1128/JVI.01905-09
Common threads in persistent viral infections
Abstract
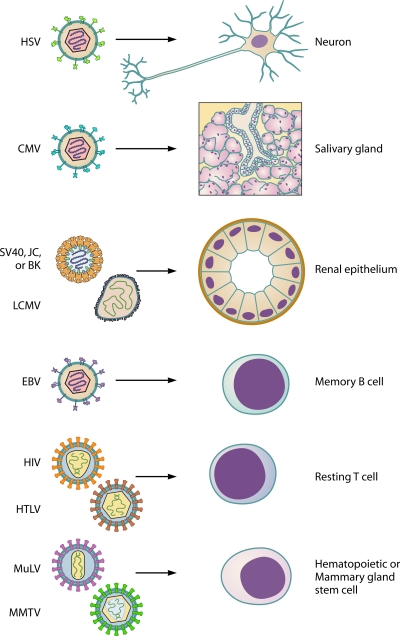
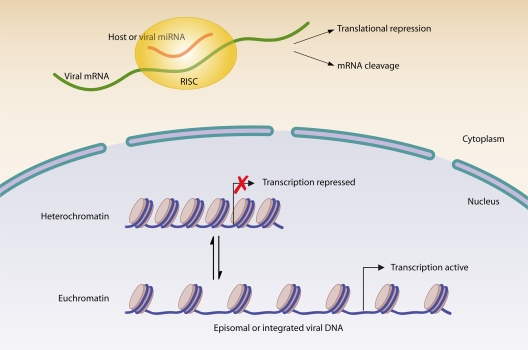
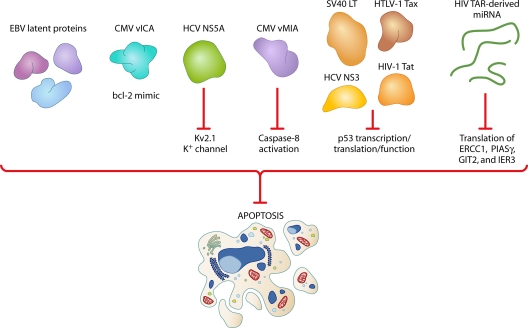
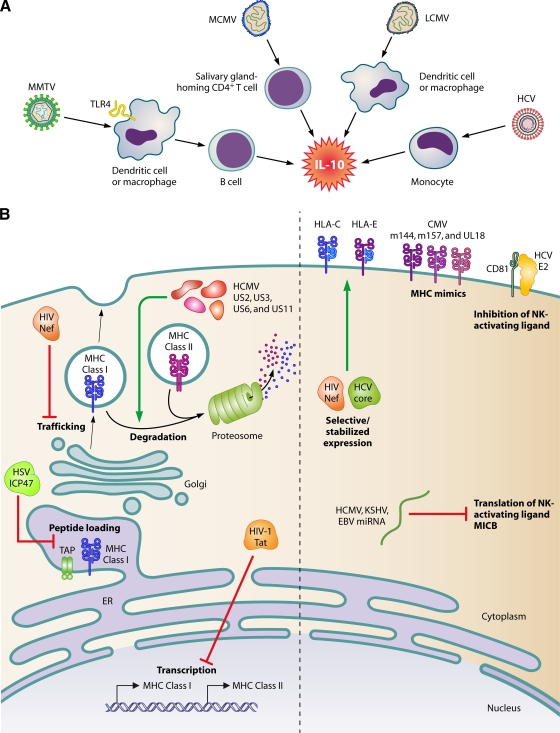
Most viral infections are self-limiting, resulting in either clearance of the pathogen or death of the host. However, a subset of viruses can establish permanent infection and persist indefinitely within the host. Even though persisting viruses are derived from various viral families with distinct replication strategies, they all utilize common mechanisms for establishment of long-lasting infections. Here, we discuss the commonalities between persistent infections with herpes-, retro-, flavi-, arena-, and polyomaviruses that distinguish them from acutely infecting viral pathogens. These shared strategies include selection of cell subsets ideal for long-term maintenance of the viral genome, modulation of viral gene expression, viral subversion of apoptotic pathways, and avoidance of clearance by the immune system.
Figures
Similar articles
-
Immunity to acute virus infections with advanced age.Curr Opin Virol. 2021 Feb;46:45-58. doi: 10.1016/j.coviro.2020.09.007. Epub 2020 Nov 4. Curr Opin Virol. 2021. PMID: 33160186 Free PMC article. Review.
-
To kill or to cure: options in host defense against viral infection.Curr Opin Immunol. 1996 Aug;8(4):478-83. doi: 10.1016/s0952-7915(96)80034-3. Curr Opin Immunol. 1996. PMID: 8794011 Review.
-
CD1-Restricted T Cells During Persistent Virus Infections: "Sympathy for the Devil".Front Immunol. 2018 Mar 19;9:545. doi: 10.3389/fimmu.2018.00545. eCollection 2018. Front Immunol. 2018. PMID: 29616036 Free PMC article. Review.
-
The role of IL-10 in regulating immunity to persistent viral infections.Curr Top Microbiol Immunol. 2011;350:39-65. doi: 10.1007/82_2010_96. Curr Top Microbiol Immunol. 2011. PMID: 20703965 Free PMC article. Review.
-
Epigenetic modulation of host: new insights into immune evasion by viruses.J Biosci. 2010 Dec;35(4):647-63. doi: 10.1007/s12038-010-0072-9. J Biosci. 2010. PMID: 21289446 Review.
Cited by
-
Metabolomic Analysis of Cricket paralysisvirus Infection in Drosophila S2 Cells Reveals Divergent Effects on Central Carbon Metabolism as Compared with Silkworm Bm5 Cells.Viruses. 2020 Apr 1;12(4):393. doi: 10.3390/v12040393. Viruses. 2020. PMID: 32244654 Free PMC article.
-
Viral manipulation of STAT3: Evade, exploit, and injure.PLoS Pathog. 2018 Mar 15;14(3):e1006839. doi: 10.1371/journal.ppat.1006839. eCollection 2018 Mar. PLoS Pathog. 2018. PMID: 29543893 Free PMC article. Review.
-
Common Themes in Zoonotic Spillover and Disease Emergence: Lessons Learned from Bat- and Rodent-Borne RNA Viruses.Viruses. 2021 Jul 31;13(8):1509. doi: 10.3390/v13081509. Viruses. 2021. PMID: 34452374 Free PMC article. Review.
-
Findings from Studies Are Congruent with Obesity Having a Viral Origin, but What about Obesity-Related NAFLD?Viruses. 2021 Jul 1;13(7):1285. doi: 10.3390/v13071285. Viruses. 2021. PMID: 34372491 Free PMC article. Review.
-
Differential Signaling and Virus Production in Calu-3 Cells and Vero Cells upon SARS-CoV-2 Infection.Biomol Ther (Seoul). 2021 May 1;29(3):273-281. doi: 10.4062/biomolther.2020.226. Biomol Ther (Seoul). 2021. PMID: 33504682 Free PMC article.
References
-
- Alcami, A. 2003. Viral mimicry of cytokines, chemokines and their receptors. Nat. Rev. Immunol. 3:36-50. - PubMed
-
- Babcock, G. J., L. L. Decker, M. Volk, and D. A. Thorley-Lawson. 1998. EBV persistence in memory B cells in vivo. Immunity 9:395-404. - PubMed
-
- Basta, S., and J. R. Bennink. 2003. A survival game of hide and seek: cytomegaloviruses and MHC class I antigen presentation pathways. Viral Immunol. 16:231-242. - PubMed
-
- Bloom, D. C. 2004. HSV LAT and neuronal survival. Int. Rev. Immunol. 23:187-198. - PubMed
-
- Bode, J. G., E. D. Brenndorfer, and D. Haussinger. 2008. Hepatitis C virus (HCV) employs multiple strategies to subvert the host innate antiviral response. Biol. Chem. 389:1283-1298. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
