

Identifying the missing steps of the autotrophic 3-hydroxypropionate CO2 fixation cycle in Chloroflexus aurantiacus
- PMID: 19955419
- PMCID: PMC2795484
- DOI: 10.1073/pnas.0908356106
Identifying the missing steps of the autotrophic 3-hydroxypropionate CO2 fixation cycle in Chloroflexus aurantiacus
Abstract
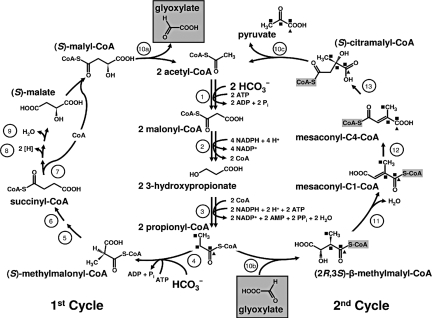
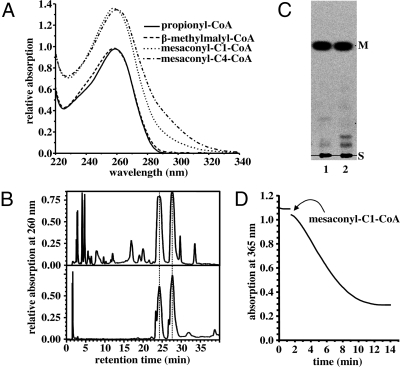
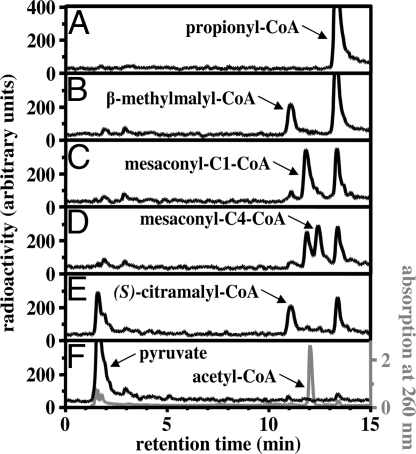
The phototrophic bacterium Chloroflexus aurantiacus uses a yet unsolved 3-hydroxypropionate cycle for autotrophic CO(2) fixation. It starts from acetyl-CoA, with acetyl-CoA and propionyl-CoA carboxylases acting as carboxylating enzymes. In a first cycle, (S)-malyl-CoA is formed from acetyl-CoA and 2 molecules of bicarbonate. (S)-Malyl-CoA cleavage releases the CO(2) fixation product glyoxylate and regenerates the starting molecule acetyl-CoA. Here we complete the missing steps devoted to glyoxylate assimilation. In a second cycle, glyoxylate is combined with propionyl-CoA, an intermediate of the first cycle, to form beta-methylmalyl-CoA. This condensation is followed by dehydration to mesaconyl-C1-CoA. An unprecedented CoA transferase catalyzes the intramolecular transfer of the CoA moiety to the C4 carboxyl group of mesaconate. Mesaconyl-C4-CoA then is hydrated by an enoyl-CoA hydratase to (S)-citramalyl-CoA. (S)-Citramalyl-CoA is cleaved into acetyl-CoA and pyruvate by a tri-functional lyase, which previously cleaved (S)-malyl-CoA and formed beta-methylmalyl-CoA. Thus, the enigmatic disproportionation of glyoxylate and propionyl-CoA into acetyl-CoA and pyruvate is solved in an elegant and economic way requiring only 3 additional enzymes. The whole bicyclic pathway results in pyruvate formation from 3 molecules of bicarbonate and involves 19 steps but only 13 enzymes. Elements of the 3-hydroxypropionate cycle may be used for the assimilation of small organic molecules. The 3-hydroxypropionate cycle is compared with the Calvin-Benson-Bassham cycle and other autotrophic pathways.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Comment in
- Proc Natl Acad Sci U S A. 106:21015.
References
-
- Thauer RK. A fifth pathway of carbon fixation. Science. 2007;318:1732–1733. - PubMed
-
- Calvin M, Bassham JA. The Photosynthesis of Carbon Compounds. New York: W. A. Benjamin, Inc.; 1962.
-
- Buchanan BB, Arnon DI. A reverse KREBS cycle in photosynthesis: Consensus at last. Photosynth Res. 1990;24:47–53. - PubMed
-
- Utter MF, Wood HG. Mechanisms of fixation of carbon dioxide by heterotrophs and autotrophs. Adv Enzymol Relat Areas Mol Biol. 1951;12:41–151. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
