

doi: 10.1097/JES.0b013e3181b7b932.
Activity-dependent plasticity of spinal locomotion: implications for sensory processing
Affiliations
- PMID: 19955866
- PMCID: PMC2790155
- DOI: 10.1097/JES.0b013e3181b7b932
Item in Clipboard
Activity-dependent plasticity of spinal locomotion: implications for sensory processing
Exerc Sport Sci Rev.
2009 Oct.
Abstract
The lumbosacral spinal cord of mammals contains the neural circuitry capable of generating full weight-bearing locomotion of the hind limbs without any supraspinal input. One or more interventions, for example, pharmacological, epidural stimulation, and/or locomotor training, however, are necessary to gain access to and modulate the properties of this circuitry and to facilitate recovery of full weight-bearing locomotion after spinal cord injury.
Figures

Correlation between mean response duration during the first 10 min of training and mRNA levels for BDNF (A), CaMKII (B), CREB (C), and synapsin I (D). There was a significant positive correlation for all the molecular systems examined except synapsin I. (Reprinted from Gómez-Pinilla F, Huie JR, Ying Z, Ferguson AR, Crown ED, Baumbauer KM, Edgerton VR, Grau JW. BDNF and learning: Evidence that instrumental training promotes learning within the spinal cord by up-regulating BDNF expression. Neuroscience. 2007; 148:893–906. Copyright © 2007 Elsevier. Used with permission.)

Instrumental learning in Non-Tr, Step-Tr, and Stand-Tr rats after 7 weeks of step training or unilateral hindlimb stand training. The flexion duration (A) and the number of flexion responses (C) for each 1 min bin over 30 min of testing are shown, as are the overall average test flexion durations (B) and response numbers (D). For flexion duration, Non-Tr and Step-Tr groups were not significantly different, while the Stand-Tr flexion duration was significantly lower relative to both the Step-Tr and Non-Tr groups (B). The average number of flexion responses was not different between groups (D). Data are mean ± SEM. *, significantly different at p< 0.05. (Reprinted from Bigbee AJ, Crown ED, Ferguson AR, Roy RR, Tillakaratne NJK, Grau, J, Edgerton VR. Two chronic motor training paradigms differentially influence acute instrumental learning in spinally transected rats. Behav. Brain Res. 2007; 180:95–101. Copyright © 2007 Elsevier. Used with permission.)

Soft robotic control schematics on how the semi-active control paradigm for step training is implemented. A moving window (thin interrupted line) bounds the desired trajectory (thick solid line) of the mouse limb during stepping. Within the window, the robotic arm allows the mouse to vary its movement. However, when the neural control desired trajectory falls outside of the window, the robot will experience a convergent velocity field, that actively returns the mouse’s limbs back within the window. This type of soft control is thought to approximate the “assist as needed” approach used by experienced therapists. (Reprinted from Cai LL, Courtine G, Fong AJ, Burdick JW, Roy RR, Edgerton VR. Plasticity of functional connectivity in the adult spinal cord. Phil. Trans. Roy. Soc. Lond. B: Biol. Sci. 2006; 361:1635–46. Copyright © 2006 Royal Society Publishing. Used with permission.)

Schematic of a semi-active fixed-trajectory paradigm for step training where the desired limb trajectory (thick solid line) is bounded by both inner and outer boundaries (thin interrupted line) is shown. The actual trajectory that the neural circuits might induce is allowed to vary within the boundary. However, once the trajectory falls outside of the boundary, the robot will actively bring it back within the boundaries. The black line with periodic dots illustrates the potential positions that the intrinsic neural control might choose to generate for any given bin time. The probability that the neural control would move the limb to the exact position defined by the thick solid line, representing a fixed trajectory, is highly unlikely. As a result, theoretically, the neural control system is continuously disrupted by the fixed trajectory paradigm. This fixed trajectory, therefore, does not allow the neural control circuitry to respond to any of its intrinsic activation patterns, but rather forces the intrinsic circuitry to continuously respond to external perturbations. This strategy for control would seem to unnecessarily disrupt the spinal circuitry and in the process minimize or even preclude the intrinsic circuitry from interpreting relevant proprioceptive information required to generate a solution (i.e., make choices) and, thus, presumably prevent the circuitry from meaningful learning phenomena. (Reprinted from Cai LL, Courtine G, Fong AJ, Burdick JW, Roy RR, Edgerton VR. Plasticity of functional connectivity in the adult spinal cord. Phil. Trans. Roy. Soc. Lond. B: Biol. Sci. 2006; 361:1635–46. Copyright © 2006 Royal Society Publishing. Used with permission.)
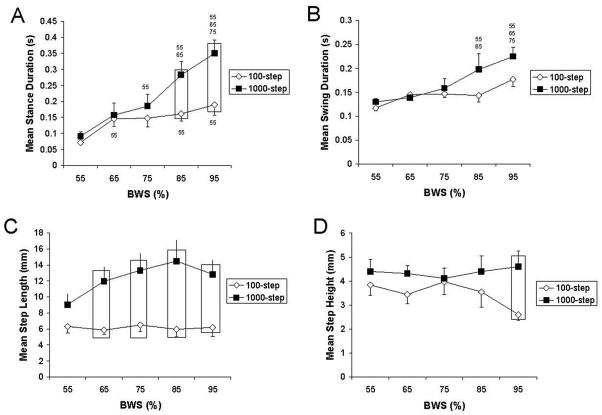
Training amount determines the responsiveness to hindlimb weight bearing. Mean stance duration (A), mean swing duration (B), mean step length (C), and mean step height (D) are shown across different levels of weight bearing for stepping tests at 8 cm · sec−1 performed after 4 weeks of training. A “55”, “65”, and “75” indicates that the data are significantly different from 55%, 65%, and 75% body weight support (BWS), respectively. For the 1000-step group in (A): the 55% BWS data are significantly different from the data at 75%, 85%, and 95% BWS; the 65% BWS data are significantly different from the data at 85% and 95% BWS; the 75% BWS data are significantly different from the data at 95% BWS. For the 100-step group in (A), the 55% BWS data are significantly different from the 65%, 85%, and 95% BWS data. For the 1000-step group in (B): the 55% and 65% BWS data are significantly different from the 85% and 95% BWS data, and the 75% BWS data are significantly different from the 95% BWS data. Boxes, significant difference between the 1000-step and 100-step groups. Values are mean ± SEM calculated from 13 rats in each group. BWS is body weight support. (Reprinted from Cha J, Heng C, Reinkensmeyer DJ, Roy RR, Edgerton VR, de Leon RD. Locomotor ability in spinal rats is dependent on the amount of activity imposed on the hindlimbs during treadmill training. J. Neurotrauma 2007; 24:1000–12. Copyright © 2007 Mary Ann Liebert, Inc. Used with permission.)
References
-
- Alvarez FJ, Taylor-Blake B, Fyffe RE, De Blas AL, Light AR. Distribution of immunoreactivity for the beta 2 and beta 3 subunits of the GABAA receptor in the mammalian spinal cord. J Comp Neurol. 1996;365:392–412. - PubMed
-
- Barbeau H, Rossignol S. Recovery of locomotion after chronic spinalization in the adult cat. Brain Res. 1987;412:84–95. - PubMed
-
- Bravo A, Bigbee AJ, Roy RR, Edgerton VR, Tobin AJ, Tillakaratne NJK. Gamma2 subunit of the GABAA receptor is increased in lumbar astrocytes in neonatally spinal cord transected rats. Soc Neurosci Abstr. 2002;28 Program No. 853.5.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
