

BK channels control cerebellar Purkinje and Golgi cell rhythmicity in vivo
- PMID: 19956720
- PMCID: PMC2776494
- DOI: 10.1371/journal.pone.0007991
BK channels control cerebellar Purkinje and Golgi cell rhythmicity in vivo
Abstract
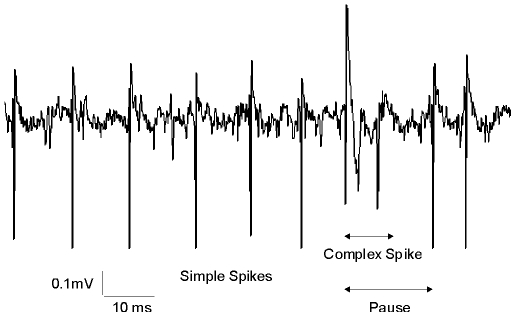
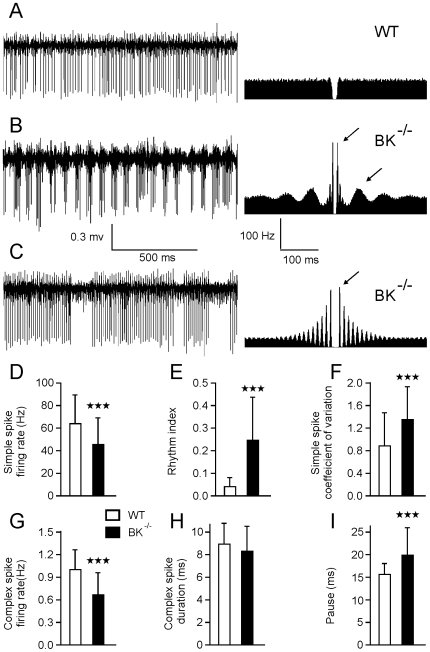
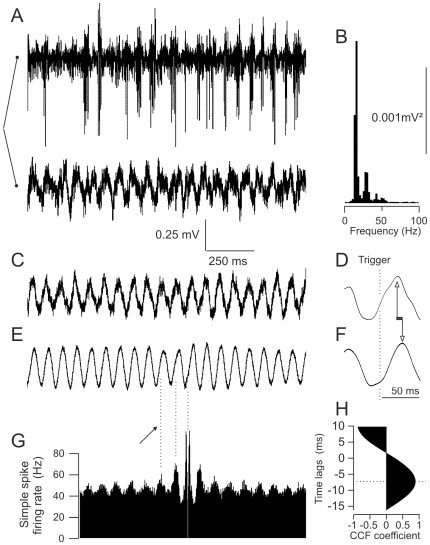
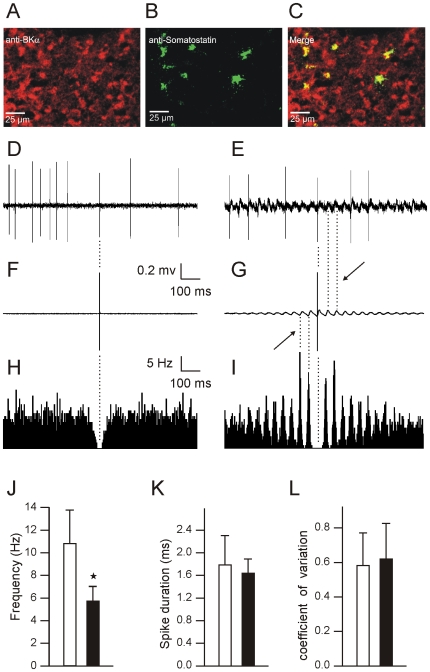
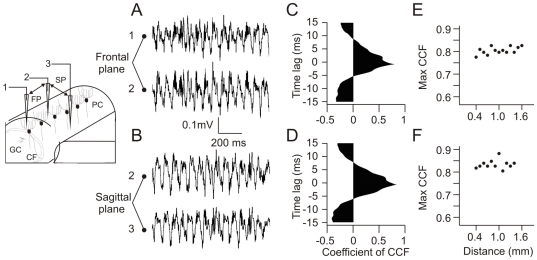
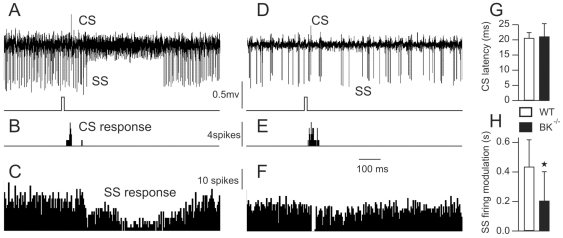
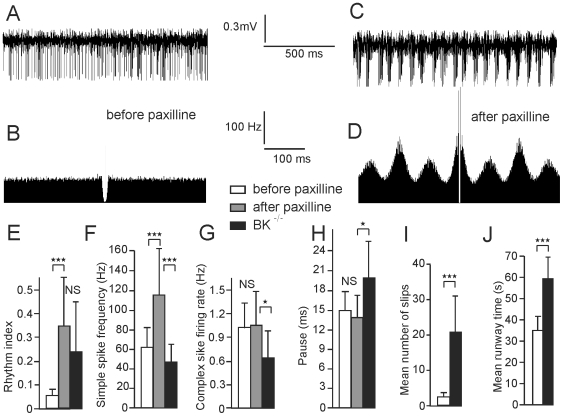
Calcium signaling plays a central role in normal CNS functioning and dysfunction. As cerebellar Purkinje cells express the major regulatory elements of calcium control and represent the sole integrative output of the cerebellar cortex, changes in neural activity- and calcium-mediated membrane properties of these cells are expected to provide important insights into both intrinsic and network physiology of the cerebellum. We studied the electrophysiological behavior of Purkinje cells in genetically engineered alert mice that do not express BK calcium-activated potassium channels and in wild-type mice with pharmacological BK inactivation. We confirmed BK expression in Purkinje cells and also demonstrated it in Golgi cells. We demonstrated that either genetic or pharmacological BK inactivation leads to ataxia and to the emergence of a beta oscillatory field potential in the cerebellar cortex. This oscillation is correlated with enhanced rhythmicity and synchronicity of both Purkinje and Golgi cells. We hypothesize that the temporal coding modification of the spike firing of both Purkinje and Golgi cells leads to the pharmacologically or genetically induced ataxia.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
