

Influence of membrane CD25 stability on T lymphocyte activity: implications for immunoregulation
- PMID: 19956753
- PMCID: PMC2775921
- DOI: 10.1371/journal.pone.0007980
Influence of membrane CD25 stability on T lymphocyte activity: implications for immunoregulation
Abstract
Background: CD25, a component of the IL-2 receptor, is important in T cell proliferation, activation induced cell death, as well as the actions of both regulatory (Treg) and effector (Teff) T cells. Recent genome wide association studies have implicated the CD25 locus as an important region for genetic susceptibility to a number of autoimmune disorders, with serum levels of soluble CD25 receptor (sCD25) serving as a potential phenotypic marker for this association. However, the functional impact of CD25 cleavage, as well as the influence of sCD25 on immunoregulatory activities, remain largely unknown and form the basis of this effort.
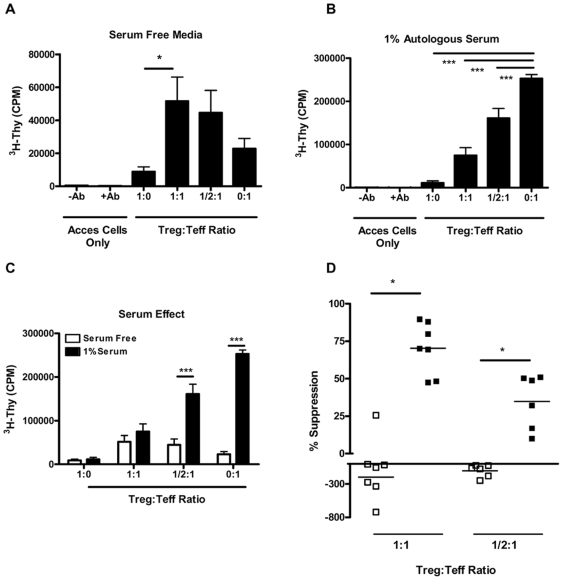
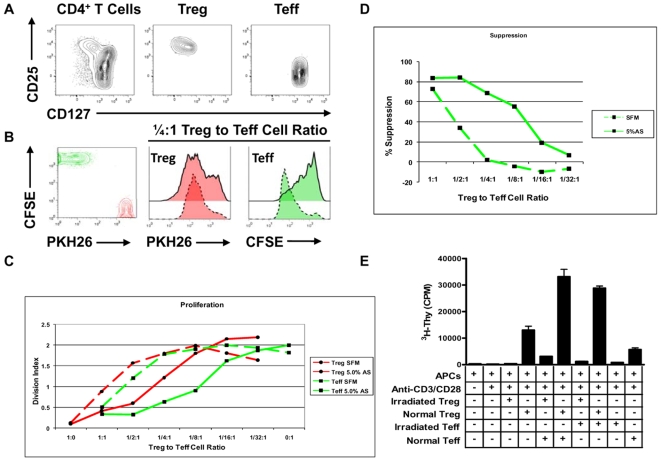
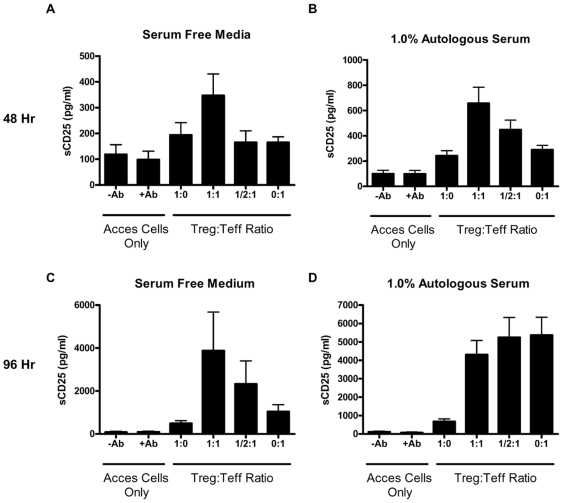
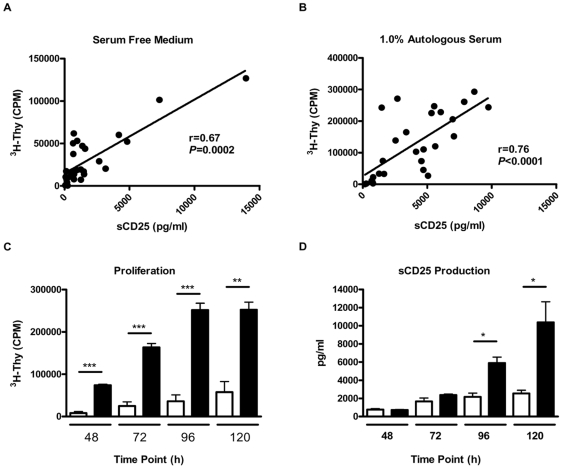
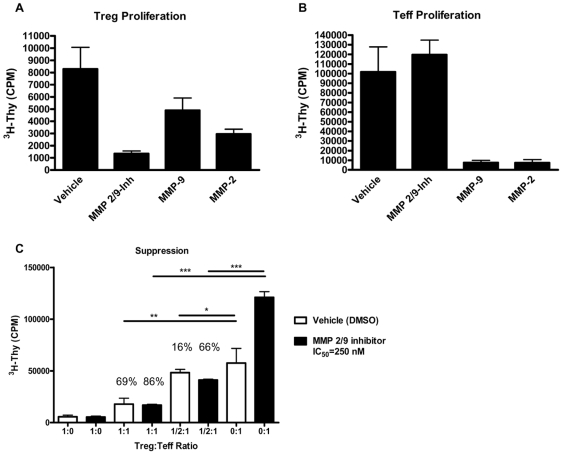
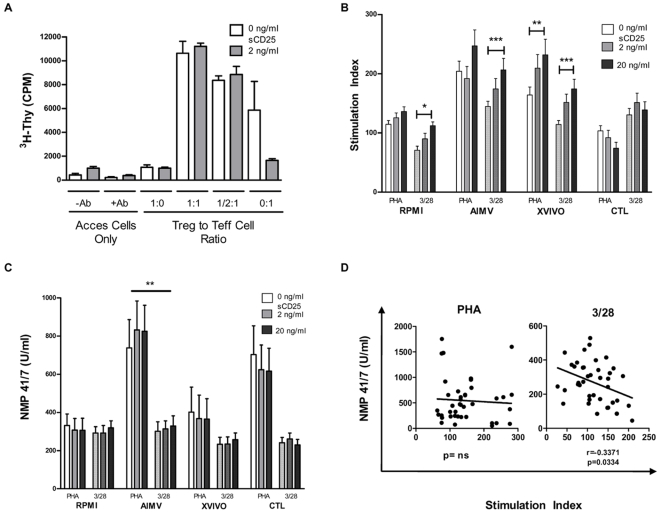
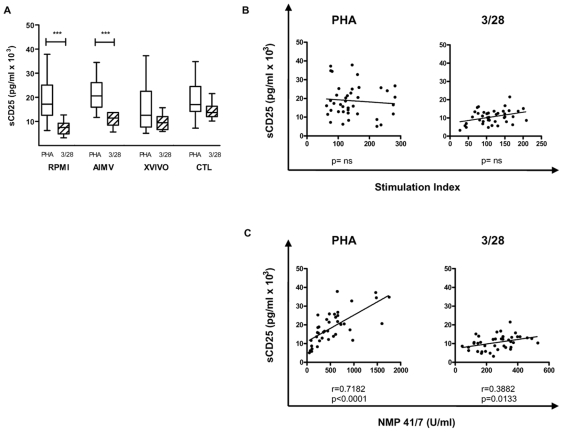
Methodology/principal findings: The generation of sCD25 by Treg (CD4(+)CD25(+)) and Teff (CD4(+)CD25(-)) cells was examined during in vitro suppression assays, efforts that demonstrated constitutive and stable surface CD25 expression on Treg throughout the period of in vitro assessment. In contrast, Teff cells increased CD25 expression during the process of in vitro suppression, with supernatant sCD25 levels correlating to the amount of cellular proliferation. Interestingly, under serum-free conditions, Tregs partially lost their characteristic anergic and suppressive properties. sCD25 supplementation at physiological concentrations to serum free in vitro suppression assays reduced Teff proliferation without specifically influencing suppression. Indeed, sCD25 production within these cultures correlated with cell death.
Conclusions/significance: These results support the notion that sCD25 functions as both a surrogate marker of T cell activation as well as an indicator of subsequent cellular death. In addition, the role of CD25 in immunomodulation is likely dependent on the local inflammatory milieu, with molecules capable of modulating surface CD25 expression playing a key role in defining immune responsiveness.
Conflict of interest statement
Figures
References
-
- Sakaguchi SS, Sakaguchi N, Asano M, Itoh M, Toda M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol. 1995;155:1151–1164. - PubMed
-
- Nelson BH, Willerford DM. Biology of the interleukin-2 receptor. Adv Immunol. 1998;70:1–81. - PubMed
-
- Smith KA. The quantal theory of immunity. Cell Res. 2006;16:11–19. - PubMed
-
- Green D R, Droin N, Pinkoski M. Activation-induced cell death in T cells. Immunol Rev. 2003;193:70–81. - PubMed
-
- Van Parijs L, Biuckians A, Ibragimov A, Alt FW, Willerford DM, et al. Functional responses and apoptosis of CD25 (IL-2R alpha)-deficient T cells expressing a transgenic antigen receptor. J Immunol. 1997;158:3738–3745. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
