

Comparative study of the gating motif and C-type inactivation in prokaryotic voltage-gated sodium channels
- PMID: 19959480
- PMCID: PMC2823509
- DOI: 10.1074/jbc.M109.057455
Comparative study of the gating motif and C-type inactivation in prokaryotic voltage-gated sodium channels
Abstract
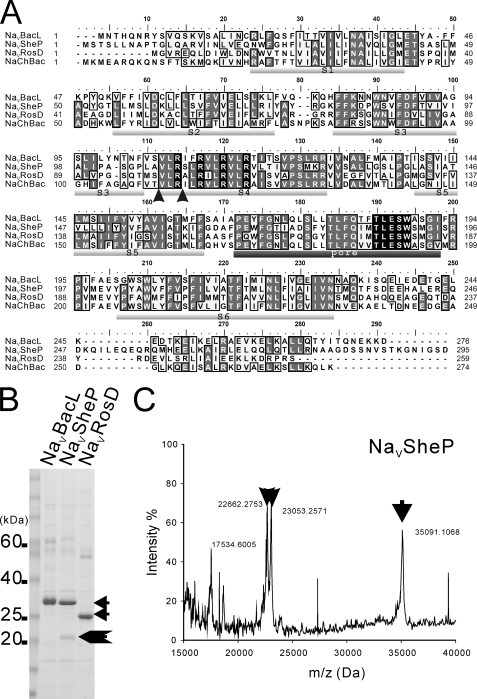
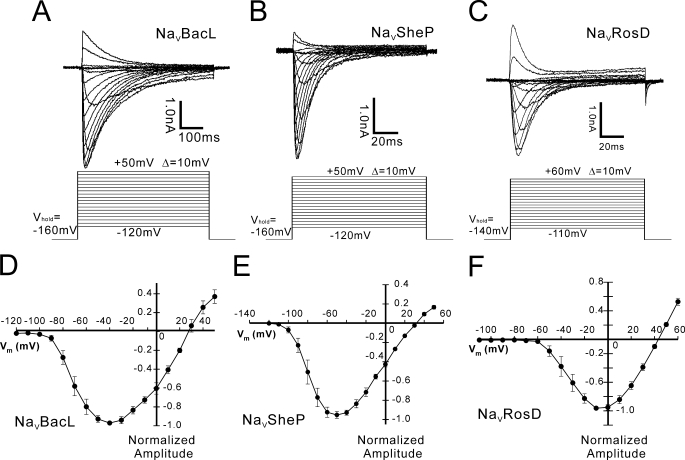
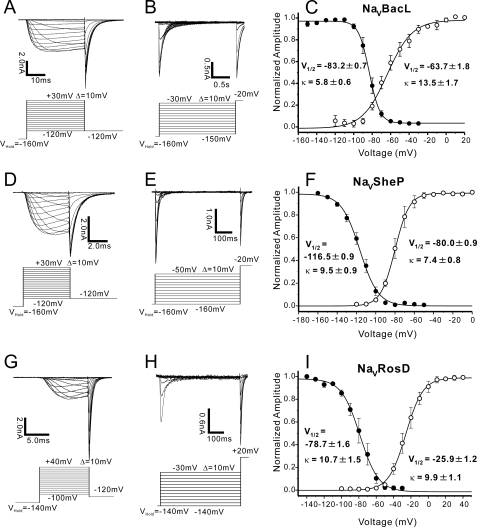
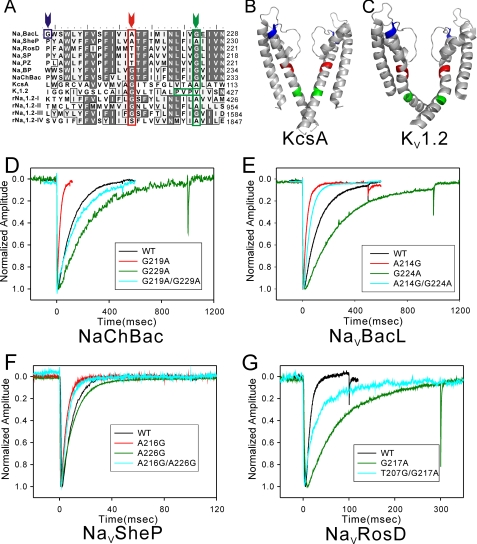
Prokaryotic voltage-gated sodium channels (Na(V)s) are homotetramers and are thought to inactivate through a single mechanism, named C-type inactivation. Here we report the voltage dependence and inactivation rate of the NaChBac channel from Bacillus halodurans, the first identified prokaryotic Na(V), as well as of three new homologues cloned from Bacillus licheniformis (Na(V)BacL), Shewanella putrefaciens (Na(V)SheP), and Roseobacter denitrificans (Na(V)RosD). We found that, although activated by a lower membrane potential, Na(V)BacL inactivates as slowly as NaChBac. Na(V)SheP and Na(V)RosD inactivate faster than NaChBac. Mutational analysis of helix S6 showed that residues corresponding to the "glycine hinge" and "PXP motif" in voltage-gated potassium channels are not obligatory for channel gating in these prokaryotic Na(V)s, but mutations in the regions changed the inactivation rates. Mutation of the region corresponding to the glycine hinge in Na(V)BacL (A214G), Na(V)SheP (A216G), and NaChBac (G219A) accelerated inactivation in these channels, whereas mutation of glycine to alanine in the lower part of helix S6 in NaChBac (G229A), Na(V)BacL (G224A), and Na(V)RosD (G217A) reduced the inactivation rate. These results imply that activation gating in prokaryotic Na(V)s does not require gating motifs and that the residues of helix S6 affect C-type inactivation rates in these channels.
Figures

Similar articles
-
Arrangement and mobility of the voltage sensor domain in prokaryotic voltage-gated sodium channels.J Biol Chem. 2011 Mar 4;286(9):7409-17. doi: 10.1074/jbc.M110.186510. Epub 2010 Dec 22. J Biol Chem. 2011. PMID: 21177850 Free PMC article.
-
Role of arginine residues on the S4 segment of the Bacillus halodurans Na+ channel in voltage-sensing.J Membr Biol. 2004 Sep 1;201(1):9-24. doi: 10.1007/s00232-004-0701-z. J Membr Biol. 2004. PMID: 15635808
-
Isoflurane inhibits NaChBac, a prokaryotic voltage-gated sodium channel.J Pharmacol Exp Ther. 2007 Sep;322(3):1076-83. doi: 10.1124/jpet.107.122929. Epub 2007 Jun 14. J Pharmacol Exp Ther. 2007. PMID: 17569823
-
Molecular basis of K+ channel inactivation gating.EXS. 1993;63:338-51. doi: 10.1007/978-3-0348-7265-2_18. EXS. 1993. PMID: 8380732 Review.
-
Evolution of voltage-gated Na(+) channels.J Exp Biol. 2002 Mar;205(Pt 5):575-84. doi: 10.1242/jeb.205.5.575. J Exp Biol. 2002. PMID: 11907047 Review.
Cited by
-
Engineered bacterial voltage-gated sodium channel platform for cardiac gene therapy.Nat Commun. 2022 Feb 2;13(1):620. doi: 10.1038/s41467-022-28251-6. Nat Commun. 2022. PMID: 35110560 Free PMC article.
-
Characterization of specific allosteric effects of the Na+ channel β1 subunit on the Nav1.4 isoform.Eur Biophys J. 2017 Jul;46(5):485-494. doi: 10.1007/s00249-016-1193-3. Epub 2016 Dec 23. Eur Biophys J. 2017. PMID: 28012039
-
Psychological stress-induced ALKBH5 deficiency promotes tumour innervation and pancreatic cancer via extracellular vesicle transfer of RNA.Nat Cell Biol. 2025 Jun;27(6):1035-1047. doi: 10.1038/s41556-025-01667-0. Epub 2025 May 26. Nat Cell Biol. 2025. PMID: 40419796
-
Generation and customization of biosynthetic excitable tissues for electrophysiological studies and cell-based therapies.Nat Protoc. 2018 May;13(5):927-945. doi: 10.1038/nprot.2018.016. Epub 2018 Apr 5. Nat Protoc. 2018. PMID: 29622805 Free PMC article.
-
Molecular basis of ion permeability in a voltage-gated sodium channel.EMBO J. 2016 Apr 15;35(8):820-30. doi: 10.15252/embj.201593285. Epub 2016 Feb 12. EMBO J. 2016. PMID: 26873592 Free PMC article.
References
-
- Hille B. (2001) Ion Channels of Excitable Membranes, 3rd Ed., Sinauer Associates, Sunderland, MA
-
- Catterall W. A. (1992) Physiol. Rev. 72, S15–S48 - PubMed
-
- Catterall W. A. (1995) Annu. Rev. Biochem. 64, 493–531 - PubMed
-
- Sato C., Ueno Y., Asai K., Takahashi K., Sato M., Engel A., Fujiyoshi Y. (2001) Nature 409, 1047–1051 - PubMed
-
- Ren D., Navarro B., Xu H., Yue L., Shi Q., Clapham D. E. (2001) Science 294, 2372–2375 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
