

Mechanisms of copy number variation and hybrid gene formation in the KIR immune gene complex
- PMID: 19959527
- PMCID: PMC2816608
- DOI: 10.1093/hmg/ddp538
Mechanisms of copy number variation and hybrid gene formation in the KIR immune gene complex
Abstract
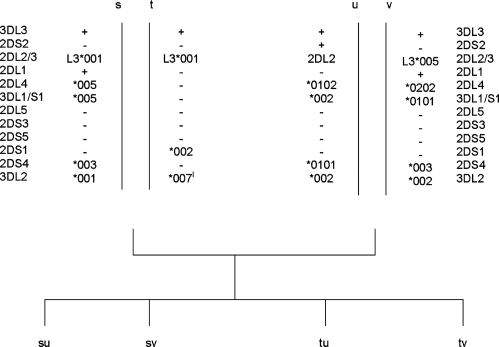
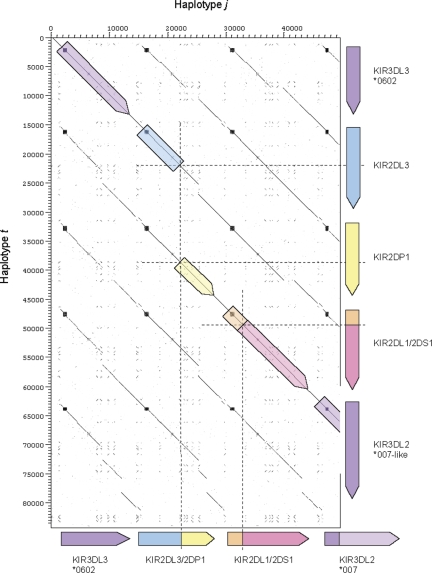
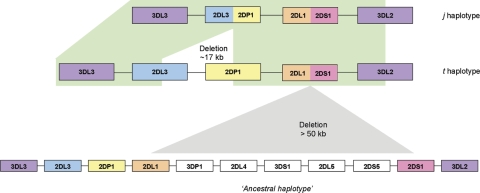
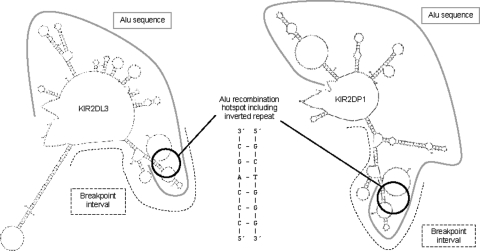
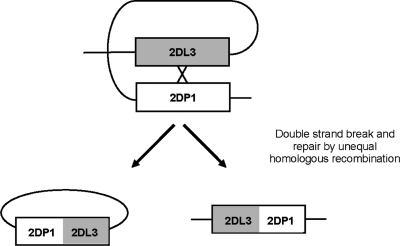
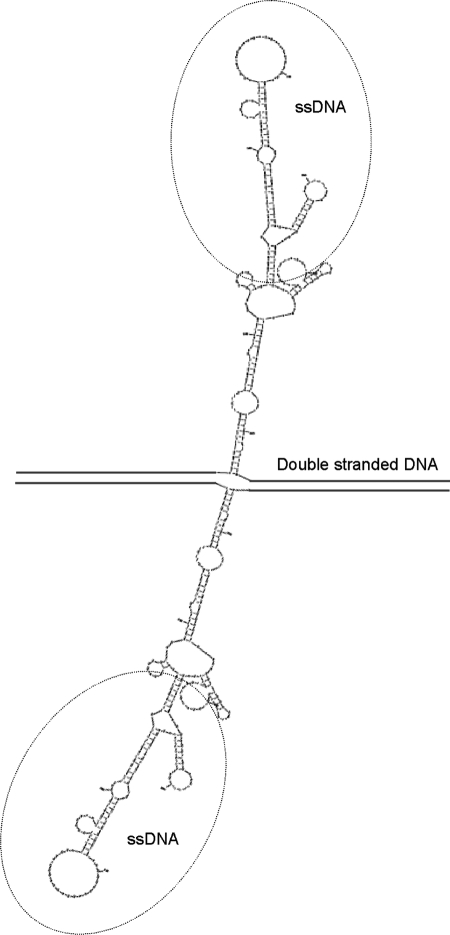
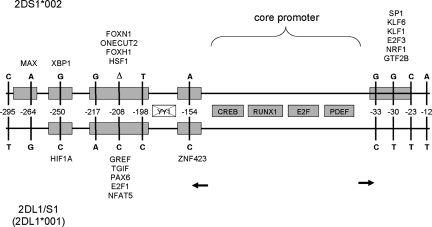
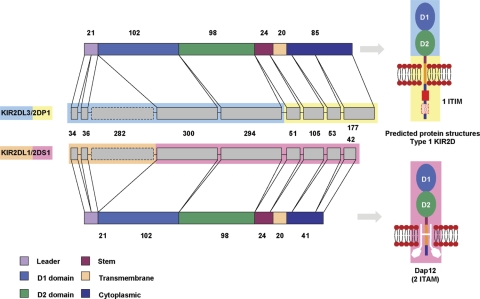
The fine-scale structure of the majority of copy number variation (CNV) regions remains unknown. The killer immunoglobulin receptor (KIR) gene complex exhibits significant CNV. The evolutionary plasticity of the KIRs and their broad biomedical relevance makes it important to understand how these immune receptors evolve. In this paper, we describe haplotype re-arrangement creating novel loci at the KIR complex. We completely sequenced, after fosmid cloning, two rare contracted haplotypes. Evidence of frequent hybrid KIR genes in samples from many populations suggested that re-arrangements may be frequent and selectively advantageous. We propose mechanisms for formation of novel hybrid KIR genes, facilitated by protrusive non-B DNA structures at transposon recombination sites. The heightened propensity to generate novel hybrid KIR receptors may provide a proactive evolutionary measure, to militate against pathogen evasion or subversion. We propose that CNV in KIR is an evolutionary strategy, which KIR typing for disease association must take into account.
Figures


Similar articles
-
Copy number variation leads to considerable diversity for B but not A haplotypes of the human KIR genes encoding NK cell receptors.Genome Res. 2012 Oct;22(10):1845-54. doi: 10.1101/gr.137976.112. Epub 2012 Sep 4. Genome Res. 2012. PMID: 22948769 Free PMC article.
-
Extensive variation in gene copy number at the killer immunoglobulin-like receptor locus in humans.PLoS One. 2013 Jun 28;8(6):e67619. doi: 10.1371/journal.pone.0067619. Print 2013. PLoS One. 2013. PMID: 23840750 Free PMC article.
-
Allele-level haplotype frequencies and pairwise linkage disequilibrium for 14 KIR loci in 506 European-American individuals.PLoS One. 2012;7(11):e47491. doi: 10.1371/journal.pone.0047491. Epub 2012 Nov 5. PLoS One. 2012. PMID: 23139747 Free PMC article.
-
Co-evolution of the MHC class I and KIR gene families in rhesus macaques: ancestry and plasticity.Immunol Rev. 2015 Sep;267(1):228-45. doi: 10.1111/imr.12313. Immunol Rev. 2015. PMID: 26284481 Free PMC article. Review.
-
The genomic context of natural killer receptor extended gene families.Immunol Rev. 2001 Jun;181:20-38. doi: 10.1034/j.1600-065x.2001.1810102.x. Immunol Rev. 2001. PMID: 11513141 Review.
Cited by
-
Local Genomic Instability of the SpTransformer Gene Family in the Purple Sea Urchin Inferred from BAC Insert Deletions.Genes (Basel). 2024 Feb 9;15(2):222. doi: 10.3390/genes15020222. Genes (Basel). 2024. PMID: 38397211 Free PMC article.
-
The role of KIR genes and their cognate HLA class I ligands in childhood acute lymphoblastic leukemia.Blood. 2014 Apr 17;123(16):2497-503. doi: 10.1182/blood-2013-11-540625. Epub 2014 Feb 11. Blood. 2014. PMID: 24518758 Free PMC article.
-
The identification of additional zebrafish DICP genes reveals haplotype variation and linkage to MHC class I genes.Immunogenetics. 2016 Apr;68(4):295-312. doi: 10.1007/s00251-016-0901-6. Epub 2016 Jan 22. Immunogenetics. 2016. PMID: 26801775 Free PMC article.
-
FcgammaRIIB in autoimmunity and infection: evolutionary and therapeutic implications.Nat Rev Immunol. 2010 May;10(5):328-43. doi: 10.1038/nri2762. Nat Rev Immunol. 2010. PMID: 20414206 Free PMC article. Review.
-
Natural killer cells and their receptors in multiple sclerosis.Brain. 2013 Sep;136(Pt 9):2657-76. doi: 10.1093/brain/aws159. Epub 2012 Jun 25. Brain. 2013. PMID: 22734127 Free PMC article. Review.
References
-
- Feuk L., Carson A.R., Scherer S.W. Structural variation in the human genome. Nat. Rev. Genet. 2006;7:85–97. - PubMed
-
- Iafrate A.J., Feuk L., Rivera M.N., Listewnik M.L., Donahoe P.K., Qi Y., Scherer S.W., Lee C. Detection of large-scale variation in the human genome. Nat. Genet. 2004;36:949–951. - PubMed
-
- Willcocks L.C., Lyons P.A., Clatworthy M.R., Robinson J.I., Yang W., Newland S.A., Plagnol V., McGovern N.N., Condliffe A.M., Chilvers E.R., et al. Copy number of FCGR3B, which is associated with systemic lupus erythematosus, correlates with protein expression and immune complex uptake. J. Exp. Med. 2008;205:1573–1582. - PMC - PubMed
-
- Yang Y., Chung E.K., Wu Y.L., Savelli S.L., Nagaraja H.N., Zhou B., Hebert M., Jones K.N., Shu Y., Kitzmiller K., et al. Gene copy-number variation and associated polymorphisms of complement component C4 in human systemic lupus erythematosus (SLE): low copy number is a risk factor for and high copy number is a protective factor against SLE susceptibility in European Americans. Am. J. Hum. Genet. 2007;80:1037–1054. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
