

Nanoridges that characterize the surface morphology of flowers require the synthesis of cutin polyester
- PMID: 19959665
- PMCID: PMC2788479
- DOI: 10.1073/pnas.0909090106
Nanoridges that characterize the surface morphology of flowers require the synthesis of cutin polyester
Abstract
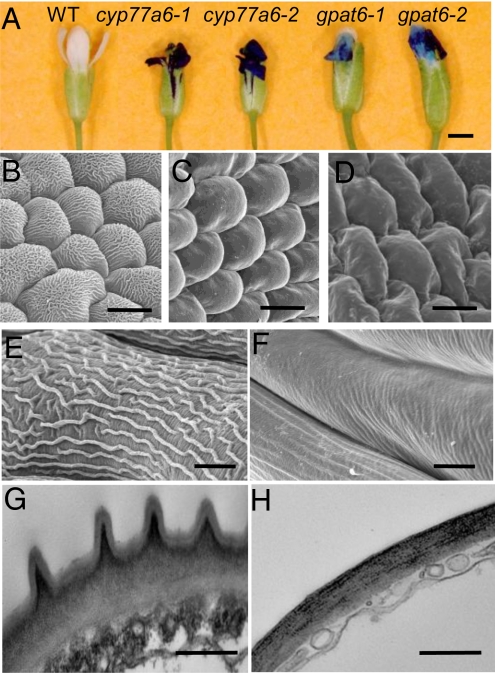
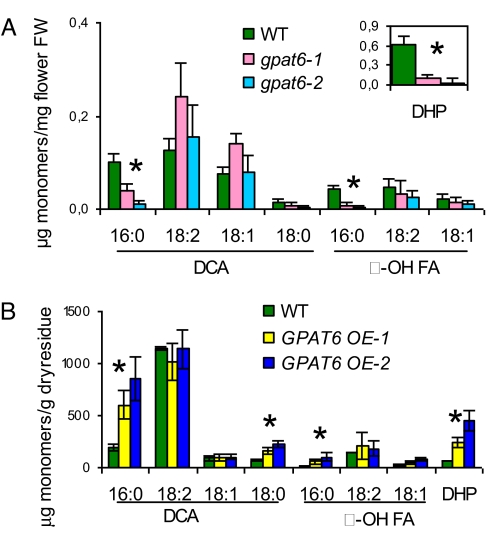
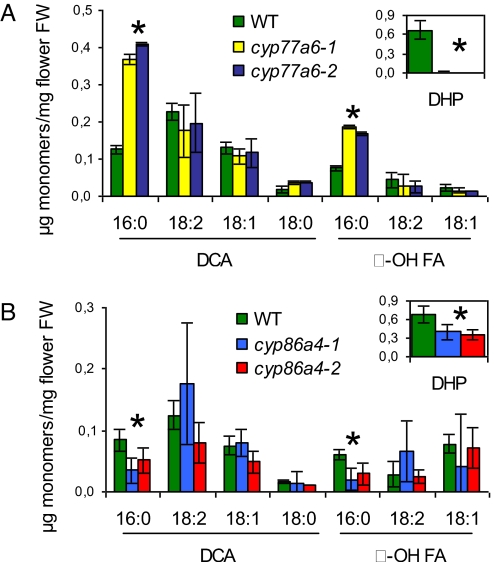
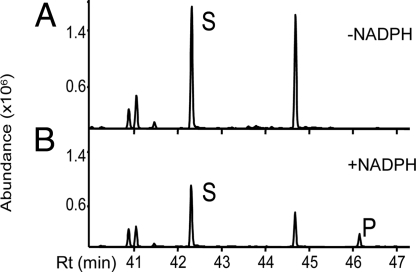
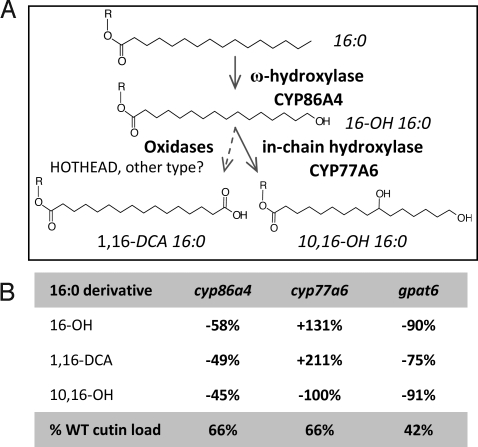
Distinctive nanoridges on the surface of flowers have puzzled plant biologists ever since their discovery over 75 years ago. Although postulated to help attract insect pollinators, the function, chemical nature, and ontogeny of these surface nanostructures remain uncertain. Studies have been hampered by the fact that no ridgeless mutants have been identified. Here, we describe two mutants lacking nanoridges and define the biosynthetic pathway for 10,16-dihydroxypalmitate, a major cutin monomer in nature. Using gene expression profiling, two candidates for the formation of floral cutin were identified in the model plant Arabidopsis thaliana: the glycerol-3-phosphate acyltransferase 6 (GPAT6) and a member of a cytochrome P450 family with unknown biological function (CYP77A6). Plants carrying null mutations in either gene produced petals with no nanoridges and no cuticle could be observed by either scanning or transmission electron microscopy. A strong reduction in cutin content was found in flowers of both mutants. In planta overexpression suggested GPAT6 preferentially uses palmitate derivatives in cutin synthesis. Comparison of cutin monomer profiles in knockouts for CYP77A6 and the fatty acid omega-hydroxylase CYP86A4 provided genetic evidence that CYP77A6 is an in-chain hydroxylase acting subsequently to CYP86A4 in the synthesis of 10,16-dihydroxypalmitate. Biochemical activity of CYP77A6 was demonstrated by production of dihydroxypalmitates from 16-hydroxypalmitate, using CYP77A6-expressing yeast microsomes. These results define the biosynthetic pathway for an abundant and widespread monomer of the cutin polyester, show that the morphology of floral surfaces depends on the synthesis of cutin, and identify target genes to investigate the function of nanoridges in flower biology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Jeffree CE. The fine structure of the plant cuticle. In: Riederer M, Müller C, editors. Biology of the Plant Cuticle. Oxford: Blackwell; 2006. pp. 11–125.
-
- Koch K, Bhushan B, Barthlott W. Diversity of structure, morphology and wetting of plant surfaces. Soft Matter. 2008;4:1943–1963.
-
- Feng L, et al. Petal effect: A superhydrophobic state with high adhesive force. Langmuir. 2008;24:4114–4119. - PubMed
-
- Barthlott W, Neinhuis C. Purity of the sacred lotus or escape from contamination in biological surfaces. Planta. 1997;202:1–8.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
