

Bactofilins, a ubiquitous class of cytoskeletal proteins mediating polar localization of a cell wall synthase in Caulobacter crescentus
- PMID: 19959992
- PMCID: PMC2824468
- DOI: 10.1038/emboj.2009.358
Bactofilins, a ubiquitous class of cytoskeletal proteins mediating polar localization of a cell wall synthase in Caulobacter crescentus
Abstract
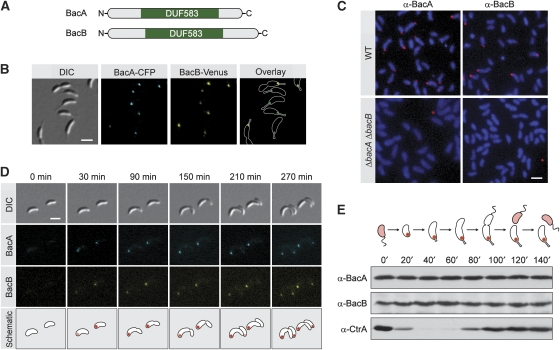
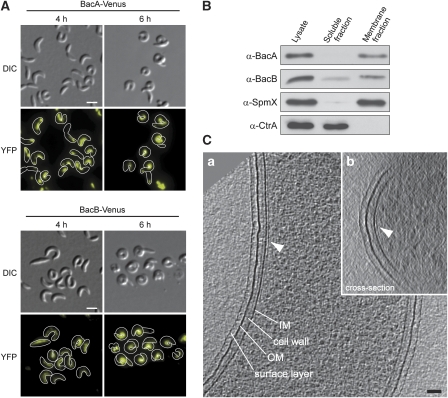
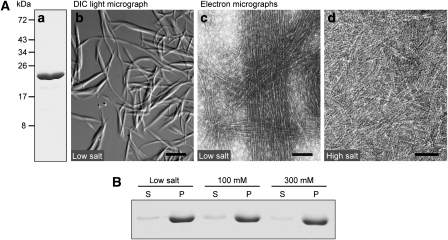
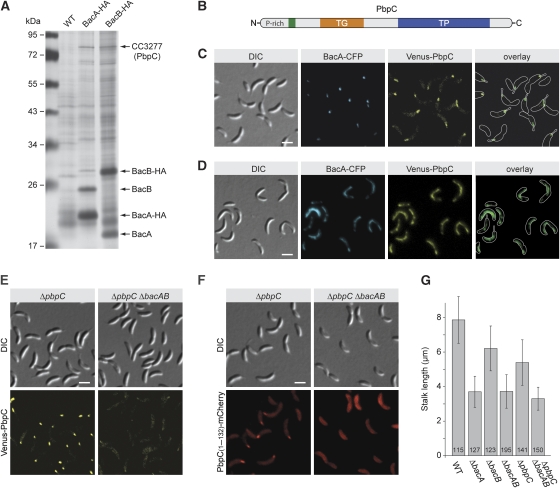
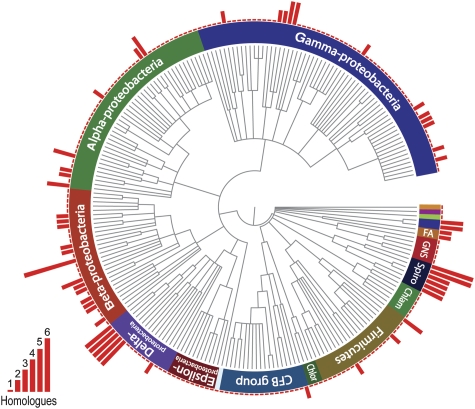
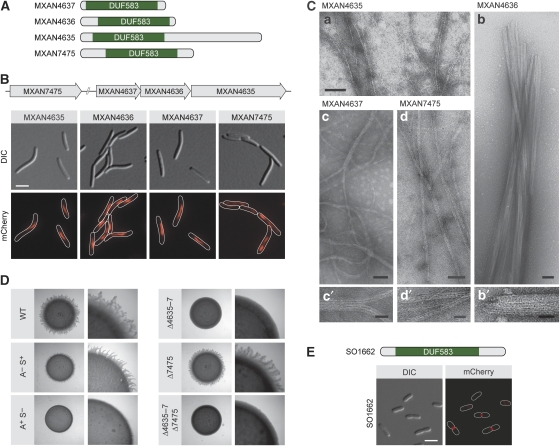
The cytoskeleton has a key function in the temporal and spatial organization of both prokaryotic and eukaryotic cells. Here, we report the identification of a new class of polymer-forming proteins, termed bactofilins, that are widely conserved among bacteria. In Caulobacter crescentus, two bactofilin paralogues cooperate to form a sheet-like structure lining the cytoplasmic membrane in proximity of the stalked cell pole. These assemblies mediate polar localization of a peptidoglycan synthase involved in stalk morphogenesis, thus complementing the function of the actin-like cytoskeleton and the cell division machinery in the regulation of cell wall biogenesis. In other bacteria, bactofilins can establish rod-shaped filaments or associate with the cell division apparatus, indicating considerable structural and functional flexibility. Bactofilins polymerize spontaneously in the absence of additional cofactors in vitro, forming stable ribbon- or rod-like filament bundles. Our results suggest that these structures have evolved as an alternative to intermediate filaments, serving as versatile molecular scaffolds in a variety of cellular pathways.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
-
- Aaron M, Charbon G, Lam H, Schwarz H, Vollmer W, Jacobs-Wagner C (2007) The tubulin homologue FtsZ contributes to cell elongation by guiding cell wall precursor synthesis in Caulobacter crescentus. Mol Microbiol 64: 938–952 - PubMed
-
- Ausmees N, Kuhn JR, Jacobs-Wagner C (2003) The bacterial cytoskeleton: an intermediate filament-like function in cell shape. Cell 115: 705–713 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
