

Pulsatility of insulin release--a clinically important phenomenon
- PMID: 19961265
- PMCID: PMC2852781
- DOI: 10.3109/03009730903366075
Pulsatility of insulin release--a clinically important phenomenon
Abstract
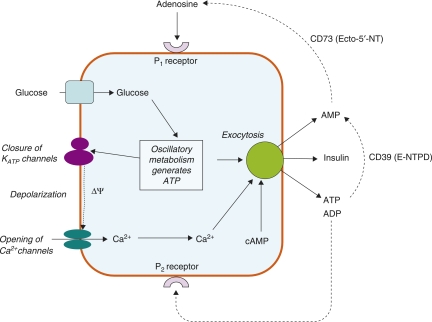
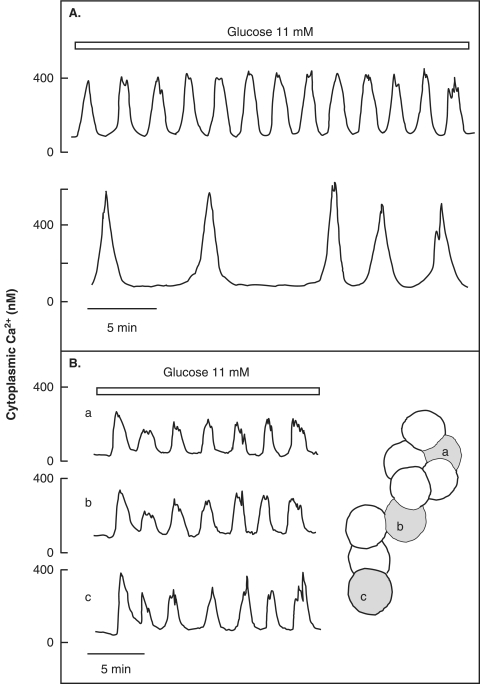
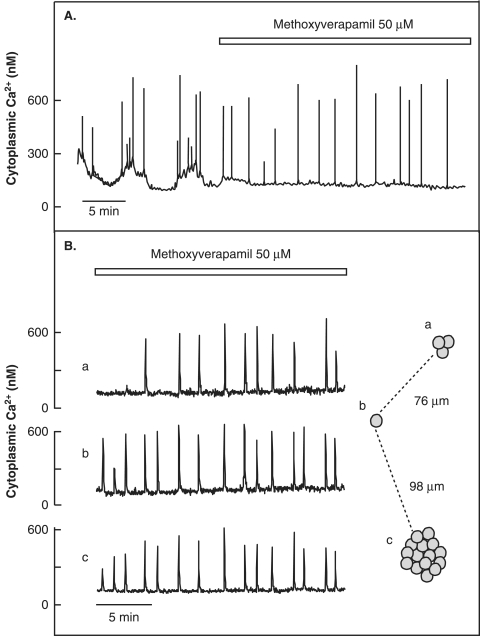
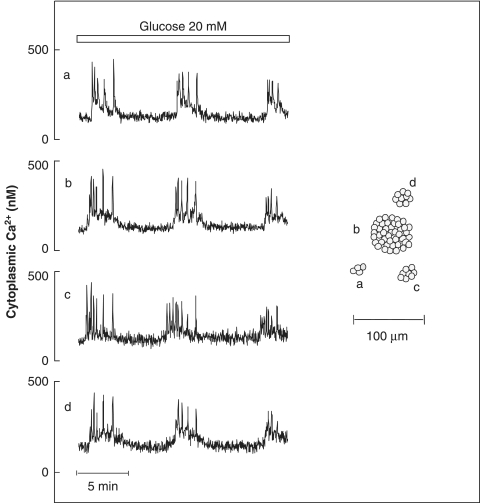
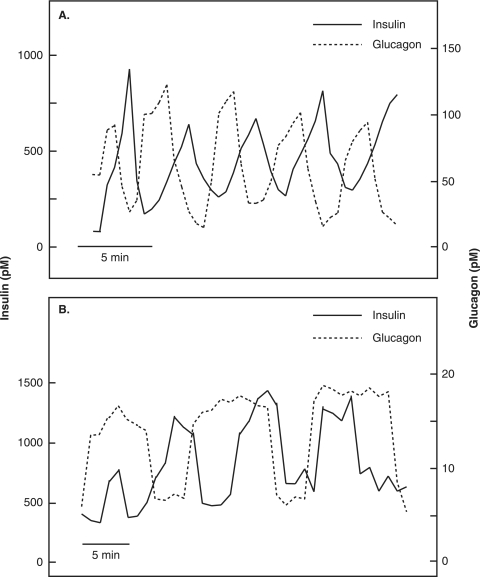
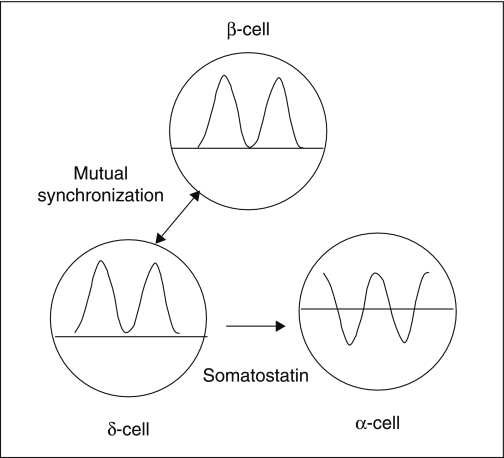
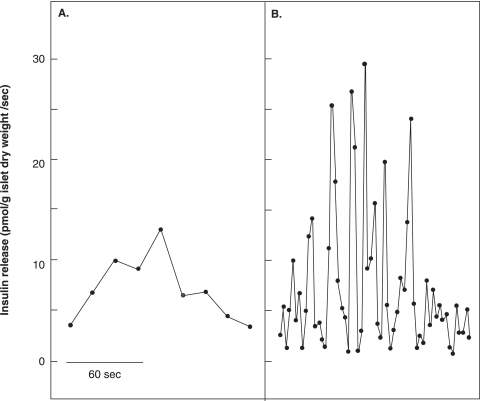
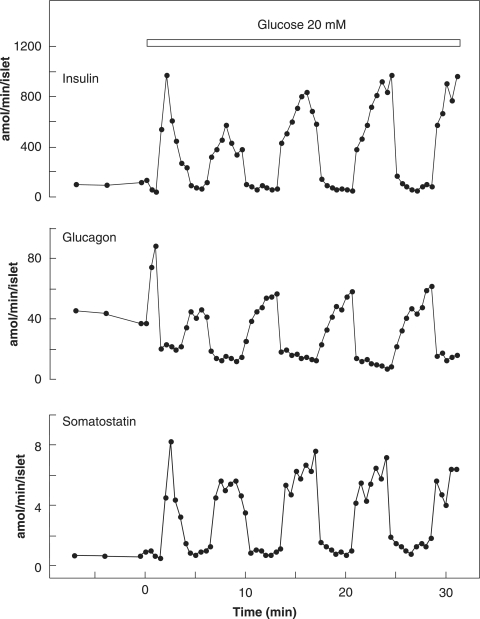
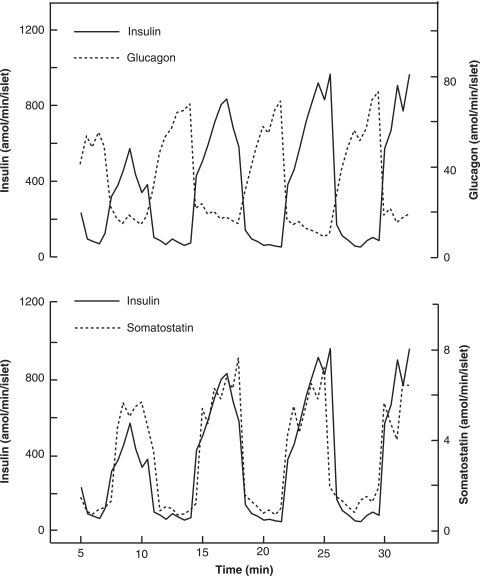
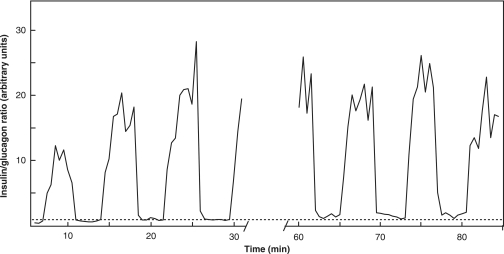
The mechanisms and clinical importance of pulsatile insulin release are presented against the background of more than half a century of companionship with the islets of Langerhans. The insulin-secreting beta-cells are oscillators with intrinsic variations of cytoplasmic ATP and Ca(2+). Within the islets the beta-cells are mutually entrained into a common rhythm by gap junctions and diffusible factors (ATP). Synchronization of the different islets in the pancreas is supposed to be due to adjustment of the oscillations to the same phase by neural output of acetylcholine and ATP. Studies of hormone secretion from the perfused pancreas of rats and mice revealed that glucose induces pulses of glucagon anti-synchronous with pulses of insulin and somatostatin. The anti-synchrony may result from a paracrine action of somatostatin on the glucagon-producing alpha-cells. Purinoceptors have a key function for pulsatile release of islet hormones. It was possible to remove the glucagon and somatostatin pulses with maintenance of those of insulin with an inhibitor of the P2Y(1) receptors. Knock-out of the adenosine A(1) receptor prolonged the pulses of glucagon and somatostatin without affecting the duration of the insulin pulses. Studies of isolated human islets indicate similar relations between pulses of insulin, glucagon, and somatostatin as found during perfusion of the rodent pancreas. The observation of reversed cycles of insulin and glucagon adds to the understanding how the islets regulate hepatic glucose production. Current protocols for pulsatile intravenous infusion therapy (PIVIT) should be modified to mimic the anti-synchrony between insulin and glucagon normally seen in the portal blood.
Figures
References
-
- Hellman B, Diderholm H. The diabetogenic effects of alloxan after elimination of extra-pancreatic factors. Acta Endocrinol (Copenh) 1955;20:81–7. - PubMed
-
- Hellman B. Actual distribution of the number and volume of the islets of Langerhans in different size classes in non-diabetic humans of varying ages. Nature. 1959;184:1498–9. - PubMed
-
- Hellman B, Sehlin J, Täljedal IB. Calcium and secretion: Distinction between two pools of glucose-sensitive calcium in pancreatic islets. Science. 1976;194:1421–3. - PubMed
-
- Hellman B. β-cell cytoplasmic Ca2+ balance as a determinant for glucose-stimulated insulin release. Diabetologia. 1985;28:494–501. - PubMed
-
- Grapengiesser E, Gylfe E, Hellman B. Three types of cytoplasmic Ca2+ oscillations in stimulated pancreatic β-cells. Arch Biochem Biophys. 1989;268:404–7. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous