

doi: 10.1111/j.1582-4934.2009.00982.x.
Epub 2009 Dec 2.
Loss of MTG16a (CBFA2T3), a novel rDNA repressor, leads to increased ribogenesis and disruption of breast acinar morphogenesis
Affiliations
- PMID: 19961547
- PMCID: PMC3828852
- DOI: 10.1111/j.1582-4934.2009.00982.x
Item in Clipboard
Loss of MTG16a (CBFA2T3), a novel rDNA repressor, leads to increased ribogenesis and disruption of breast acinar morphogenesis
J Cell Mol Med.
2010 Jun.
Erratum in
- J Cell Mol Med. 2010 Aug;14(8):2186
Abstract
Human MTG16a (CBFA2T3), a chromatin repressor with nucleolar localization, was described to act as a suppressor of breast tumourigenesis. Here we show that MTG16a is a novel ribosomal gene repressor, which can counteract MYC-driven activation of ribosomal RNA (rRNA) transcription. We also show that either knocking down MTG16a by RNA interference, or sequestering MTG16a outside the nucleolus of human breast epithelial cells, hampers acinar morphogenesis concomitant with up-regulation of rRNA synthesis and increased ribogenesis. This is the first demonstration that loss of MTG16a function in the nucleolus of breast epithelial cells can induce morphological and molecular changes typical of breast cancer initiation.
Figures

MTG16a localizes in the nucleolus and represses rRNA synthesis in Hela cells. (A) Immunocytochemistry showing that exogenous MTG16a-V5 localizes mainly in the nucleolus of Hela cells. (B) ChIP with anti-MTG16 of untransfected Hela, followed by real time PCR with primers amplifying either a region in the rDNA promoter or a control region in the 28S rDNA (regions ‘a’ and ‘c’, respectively, in the scheme on top), shows that endogenous MTG16 is associated significantly (P < 0.01) more with the rDNA promoter than the control region (bottom, left). ChIP with both anti-MTG16 and anti-V5 of Hela transiently transfected with MTG16a-V5 shows that exogenous MTG16a is associated significantly (P < 0.05) more with the rDNA promoter than the control region (bottom, right). (C) [3H]Uridine incorporation analysis shows that transient expression of MTG16a-V5 significantly (P < 0.001) decreases rRNA synthesis in Hela cells; the right panel shows that equal amounts of RNA were loaded on agarose gel for quantification of the [3H]-labelled rRNA (left). qRT-PCR with primers amplifying a region in the 5′ ETS of pre-rRNA (region ‘b’) shows that transient expression of MTG16a-V5 significantly (P < 0.001) decreases the rRNA level in Hela cells (right). (D) ChIP analysis shows that transient expression of MTG16a-V5 significantly (P < 0.01) decreases H4 acetylation (AcH4, left) and significantly (P < 0.01) increases histone H3 lysine 9 tri-methylation (tri-met-H3K9, right) at the rDNA promoter. The bars in this figure represent the average of three determinations ± standard deviation.

MTG16a counteracts MYC-induced up-regulation of rRNA synthesis in human fibroblasts. (A) [3H]Uridine incorporation (left) and qRT-PCR analysis (right) show that human fibroblasts stably expressing exogenous MYC (HFF-MYC) display significantly (P < 0.05) higher rRNA synthesis and rRNA level relative to control fibroblasts carrying the cognate empty vector (HFF-ctrl). (B) Immunocytochemistry with anti-V5 antibody shows that MTG16a-V5 transiently transfected into HFF-MYC cells can localize in the nucleolus. (C) Transient transfection of HFF-MYC with MTG16a-V5 leads to a significant (P < 0.05) decrease of both rRNA synthesis (left) and rRNA level (right), as shown by [3H]Uridine incorporation and qRT-PCR, respectively. The bars in this figure represent the average of three determinations ± standard deviation.

MTG16a down-regulates rRNA synthesis in human breast epithelial cells. (A) RT-PCR showing that the two human breast epithelial cell lines MCF10A and HME1 express only MTG16a but not MTG16b (top); Western blot of nuclear extracts using anti-MTG16 antibody confirms MTG16a expression in both cell lines (bottom). (B) Immuncytochemistry showing that endogenous MTG16a, detected with anti-MTG16 (green), can localize both in the nucleoplasm and in the B23-positive nucleoli (red) of both HME1 and MCF10A. Nuclei are counterstained with DAPI (blue). (C) Exogenous MTG16a-V5 can be detected both in the nucleoplasm and the nucleolus by IC with anti-V5 antibody. (D) Transient expression of MTG16a-V5 in HME1 cells induces a significant (P < 0.05) decrease of both rRNA synthesis (left) and rRNA level (right). The bars in this figure represent the average of three determinations ± standard deviation.

MTG16a counteracts MYC-induced up-regulation of rRNA synthesis in human breast epithelial cells. (A) Immunocytochemistry with anti-MYC antibody (left) followed by digital quantification (right) shows MYC overexpression in HME1 cells stably transfected with human MYC (HME1-MYC) relative to control HME1 cells transfected with the cognate empty vector. (B) HME1-MYC cells display significant (P < 0.05) up-regulation of both rRNA synthesis (left) and rRNA level (right) relative to control cells carrying the cognate empty vector (HME1-ctrl). (C) Transient expression of exogenous MTG16a-V5 significantly (P < 0.05) decreases both rRNA synthesis (left) and rRNA level (right) of HME1-MYC cells. The bars in this figure represent the average of three determinations ± standard deviation.
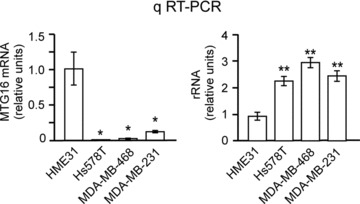
Low MTG16 expression is associated with an increased rRNA level in breast cancer cells. qRT-PCR showing that the breast cancer cell lines Hs578T, MDA-MB-231 and MDA-MB-468 have a significantly (P < 0.05) lower level of MTG16 (left) and a significantly (P < 0.01) higher rRNA level (right) than the primary breast epithelial cell line HME31. The bars in this figure represent the average of three determinations ± standard deviation.

MTG16a knockdown in human breast epithelial cells induces an increase of rRNA synthesis and NORs. (A) Exogenous MTG16a-GFP expression in HME1 cells (left) decreases significantly after RNA interference with shMTG16-1 and shMTG16-2 sequences (right). (B) HME1 clones carrying either pSuper-MTG16-1 (such as H16-1a and H16-1b) or pSuper-MTG16-2 (such as H16-2a and H16-2b) have a decreased MTG16a level relative to the HME1 parental cell line or a control clone (H-ctrl), as shown by Western blot with anti-MTG16 on nuclear extracts. Histone H3 was used as loading control. (C) HME1-derived MTG16a knockdown clones display both a significantly (P < 0.01) higher rRNA synthesis (left) and a significantly (P < 0.05) higher rRNA level (right) relative to control cells. The bars represent the average of at least three determinations on two independent clones ± standard deviation. (D) Digital analysis of silver stained nucleoli (right) shows a significant increase both in NORs number and NORs total area in HME1-derived MTG16a knockdown clones relative to the cognate control clones (left).
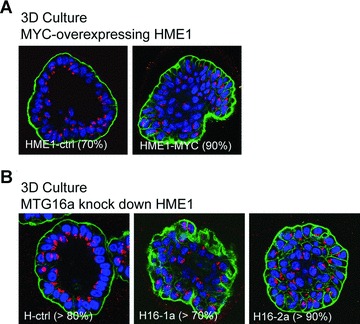
Up-regulation of rRNA synthesis in breast epithelial cells is associated with impaired acinar morphogenesis. (A) Three-dimensional (3D) culture on reconstituted basement membrane followed by confocal microscopy shows that HME1-MYC cells are unable of proper acinar morphogenesis in more than 90% of the acini, as indicated by the inability to form hollow, polarized acini, with organized arrangement of the nuclei (blue), presence of an outer layer of integrin (green) and apical localization of the Golgi apparatus (red). (B) HME1 clones (H16-1a and H16-2a) knocked down for MTG16a are also unable of proper acinar morphogenesis in 3D cultures in more than 70% of the acini, as opposed to control clones forming more than 80% hollow, polarized acini.
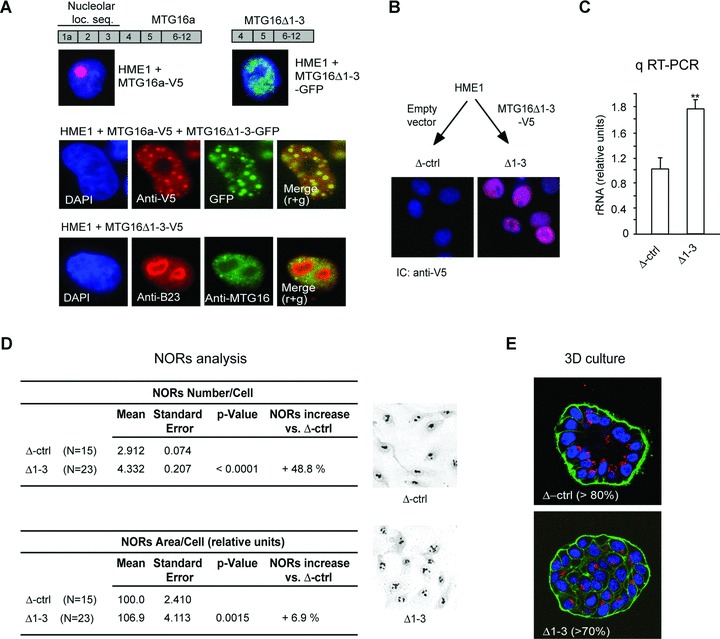
Sequestering MTG16a outside the nucleolus is sufficient to increase rRNA level and ribogenesis as well as disrupt acinar morphogenesis. (A) The V5-tagged wild-type MTG16a protein, containing the nucleolar localization sequence encoded by exons 1a-3, is targeted to the nucleolus of HME1 cells, while the GFP-tagged MTG16Δ1-3 deletion mutant protein, lacking the nucleolar localization sequence, cannot localize in the nucleolus (top). Immunocytochemistry of HME1 cells co-transfected with MTG16a-V5 and MTG16Δ1-3-GFP shows co-localization of the two exogenous proteins in the nucleoplasm (middle). Expression of MTG16Δ1-3-V5 in HME1 cells leads to depletion of endogenous MTG16a (detected with anti-MTG16, green) from the nucleoli (visualized with anti-B23, red). Nuclei are counterstained with DAPI (blue) (bottom). (B) Representative HME1 clone stably transfected with MTG16Δ1-3-V5 showing localization of the deletion mutant protein (red) in the nucleus (blue). (C) Δ1-3 cells display a significantly (P < 0.01) higher rRNA level than control cells (Δctrl). (D) Digital analysis of silver stained slides (right) shows a significant increase both in NORs number and NORs total area in Δ1-3 cells when compared to their cognate control cells. (E) Δ1-3 cells are unable of proper acinar morphogenesis when grown in 3D basement membrane culture.
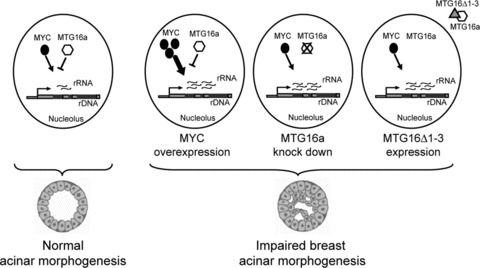
Scheme summarizing the potential relationship between rRNA synthesis level and breast acinar morphogenesis. A proper rRNA synthesis level, maintained by the opposing action of activator complexes (e.g. containing MYC) and repressor complexes (e.g. containing MTG16a), would associate with normal acinar morphogenesis (left). An increased rRNA synthesis level, due to different factors (e.g. MYC overexpression, MTG16a knockdown, or depletion of MTG16a from the nucleolus via the MTG16Δ1-3 mutant) able to alter the balance between rDNA activator and repressor complexes, would associate with impaired acinar morphogenesis (right).
Similar articles
-
The transcriptional corepressor MTG16a contains a novel nucleolar targeting sequence deranged in t (16; 21)-positive myeloid malignancies.Oncogene. 2002 Sep 26;21(43):6703-12. doi: 10.1038/sj.onc.1205882. Oncogene. 2002. PMID: 12242670
-
Che-1/AATF binds to RNA polymerase I machinery and sustains ribosomal RNA gene transcription.Nucleic Acids Res. 2020 Jun 19;48(11):5891-5906. doi: 10.1093/nar/gkaa344. Nucleic Acids Res. 2020. PMID: 32421830 Free PMC article.
-
DEAR1 is a dominant regulator of acinar morphogenesis and an independent predictor of local recurrence-free survival in early-onset breast cancer.PLoS Med. 2009 May 26;6(5):e1000068. doi: 10.1371/journal.pmed.1000068. Epub 2009 May 5. PLoS Med. 2009. PMID: 19536326 Free PMC article.
-
Ribosomal RNA Transcription Regulation in Breast Cancer.Genes (Basel). 2021 Mar 29;12(4):502. doi: 10.3390/genes12040502. Genes (Basel). 2021. PMID: 33805424 Free PMC article. Review.
-
The multifunctional nucleolus.Nat Rev Mol Cell Biol. 2007 Jul;8(7):574-85. doi: 10.1038/nrm2184. Nat Rev Mol Cell Biol. 2007. PMID: 17519961 Review.
Cited by
-
Undermining ribosomal RNA transcription in both the nucleolus and mitochondrion: an offbeat approach to target MYC-driven cancer.Oncotarget. 2017 Dec 22;9(4):5016-5031. doi: 10.18632/oncotarget.23579. eCollection 2018 Jan 12. Oncotarget. 2017. PMID: 29435159 Free PMC article.
-
Mammary epithelial morphogenesis and early breast cancer. Evidence of involvement of basal components of the RNA Polymerase I transcription machinery.Cell Cycle. 2016 Sep 16;15(18):2515-26. doi: 10.1080/15384101.2016.1215385. Epub 2016 Aug 2. Cell Cycle. 2016. PMID: 27485818 Free PMC article.
-
3D Mammary Epithelial Cell Models: A Goldmine of DCIS Biomarkers and Morphogenetic Mechanisms.Cancers (Basel). 2019 Jan 23;11(2):130. doi: 10.3390/cancers11020130. Cancers (Basel). 2019. PMID: 30678048 Free PMC article.
-
The emerging role of RNA polymerase I transcription machinery in human malignancy: a clinical perspective.Onco Targets Ther. 2013 Jul 19;6:909-16. doi: 10.2147/OTT.S36627. Print 2013. Onco Targets Ther. 2013. PMID: 23888116 Free PMC article.
-
Tracing anti-cancer and cancer-promoting actions of all-trans retinoic acid in breast cancer to a RARα epigenetic mechanism of mammary epithelial cell fate.Oncotarget. 2016 Dec 27;7(52):87064-87080. doi: 10.18632/oncotarget.13500. Oncotarget. 2016. PMID: 27894085 Free PMC article.
References
-
- Boisvert FM, van Koningsbruggen S, Navascues J, et al. The multifunctional nucleolus. Nat Rev Mol Cell Biol. 2007;8:574–85. - PubMed
-
- Grummt I. Life on a planet of its own: regulation of RNA polymerase I transcription in the nucleolus. Genes Dev. 2003;17:1691–702. - PubMed
-
- Grummt I. Different epigenetic layers engage in complex crosstalk to define the epigenetic state of mammalian rRNA genes. Hum Mol Genet. 2007:R21–7. 16 Spec No 1. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
