

O-GlcNAc protein modification in plants: Evolution and function
- PMID: 19961900
- PMCID: PMC2815191
- DOI: 10.1016/j.bbagen.2009.11.016
O-GlcNAc protein modification in plants: Evolution and function
Abstract
The role in plants of posttranslational modification of proteins with O-linked N-acetylglucosamine and the evolution and function of O-GlcNAc transferases responsible for this modification are reviewed. Phylogenetic analysis of eukaryotic O-GlcNAc transferases (OGTs) leads us to propose that plants have two distinct OGTs, SEC- and SPY-like, that originated in prokaryotes. Animals and some fungi have a SEC-like enzyme while plants have both. Green algae and some members of the Apicomplexa and amoebozoa have the SPY-like enzyme. Interestingly the progenitor of the Apicomplexa lineage likely had a photosynthetic plastid that persists in a degenerated form in some species, raising the possibility that plant SPY-like OGTs are derived from a photosynthetic endosymbiont. OGTs have multiple tetratricopeptide repeats (TPRs) that within the SEC- and SPY-like classes exhibit evidence of strong selective pressure on specific repeats, suggesting that the function of these repeats is conserved. SPY-like and SEC-like OGTs have both unique and overlapping roles in the plant. The phenotypes of sec and spy single and double mutants indicate that O-GlcNAc modification is essential and that it affects diverse plant processes including response to hormones and environmental signals, circadian rhythms, development, intercellular transport and virus infection. The mechanistic details of how O-GlcNAc modification affects these processes are largely unknown. A major impediment to understanding this is the lack of knowledge of the identities of the modified proteins.
Copyright 2009 Elsevier B.V. All rights reserved.
Figures
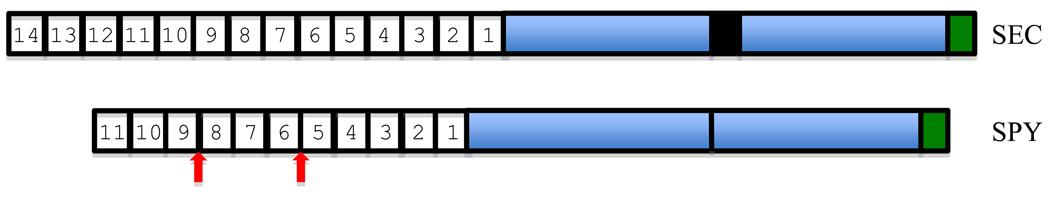
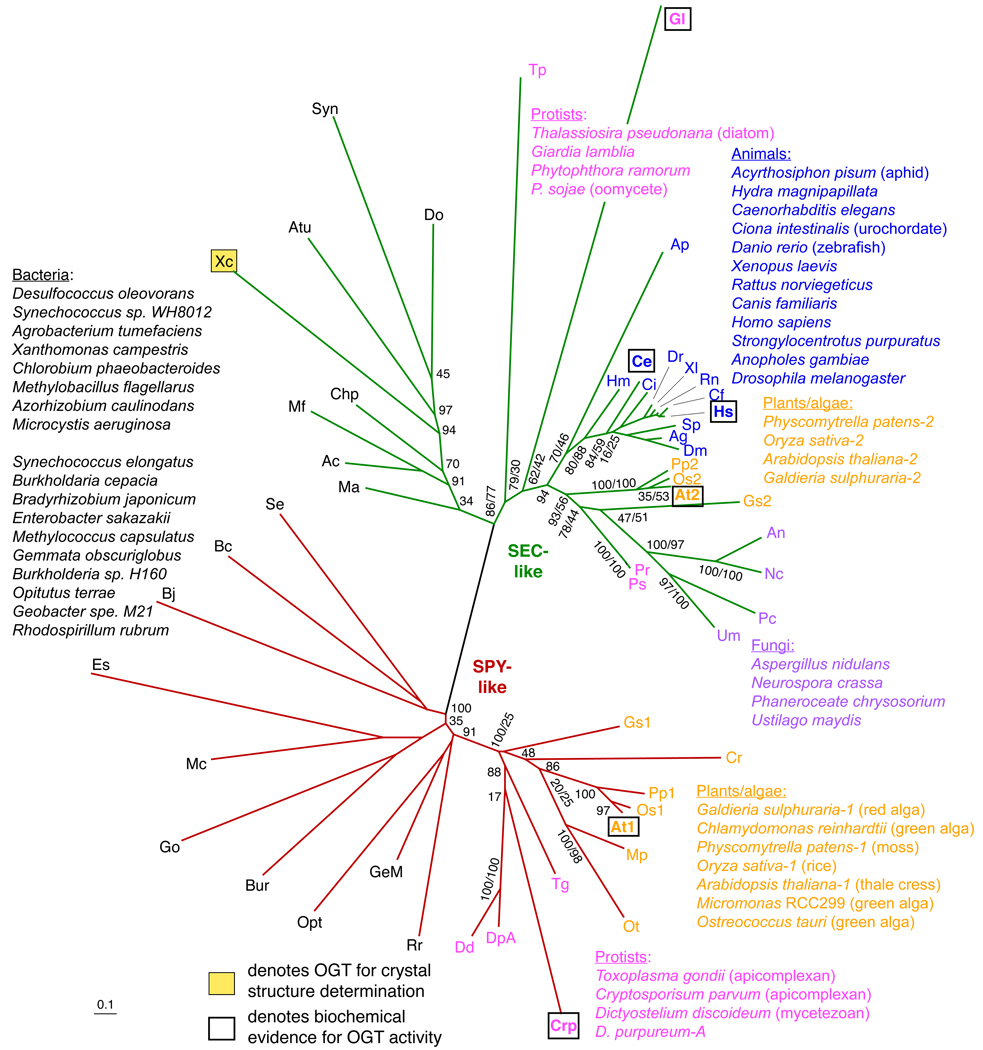
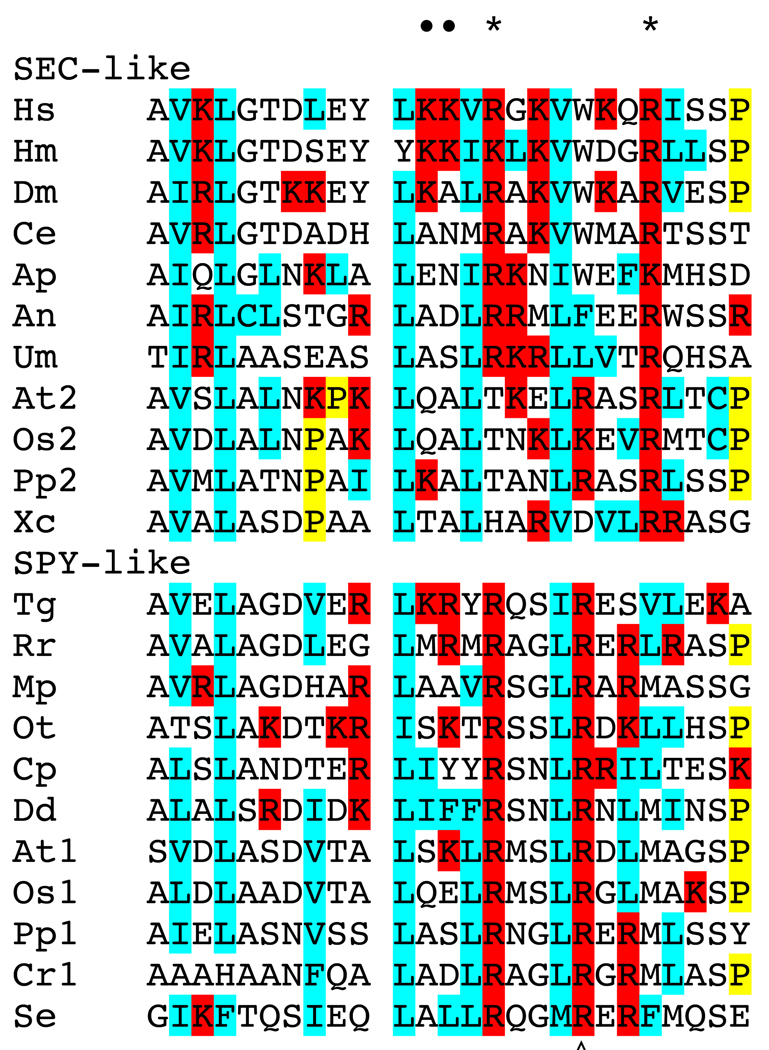
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
