

The pivotal role of the complement system in aging and age-related macular degeneration: hypothesis re-visited
- PMID: 19961953
- PMCID: PMC3641842
- DOI: 10.1016/j.preteyeres.2009.11.003
The pivotal role of the complement system in aging and age-related macular degeneration: hypothesis re-visited
Abstract
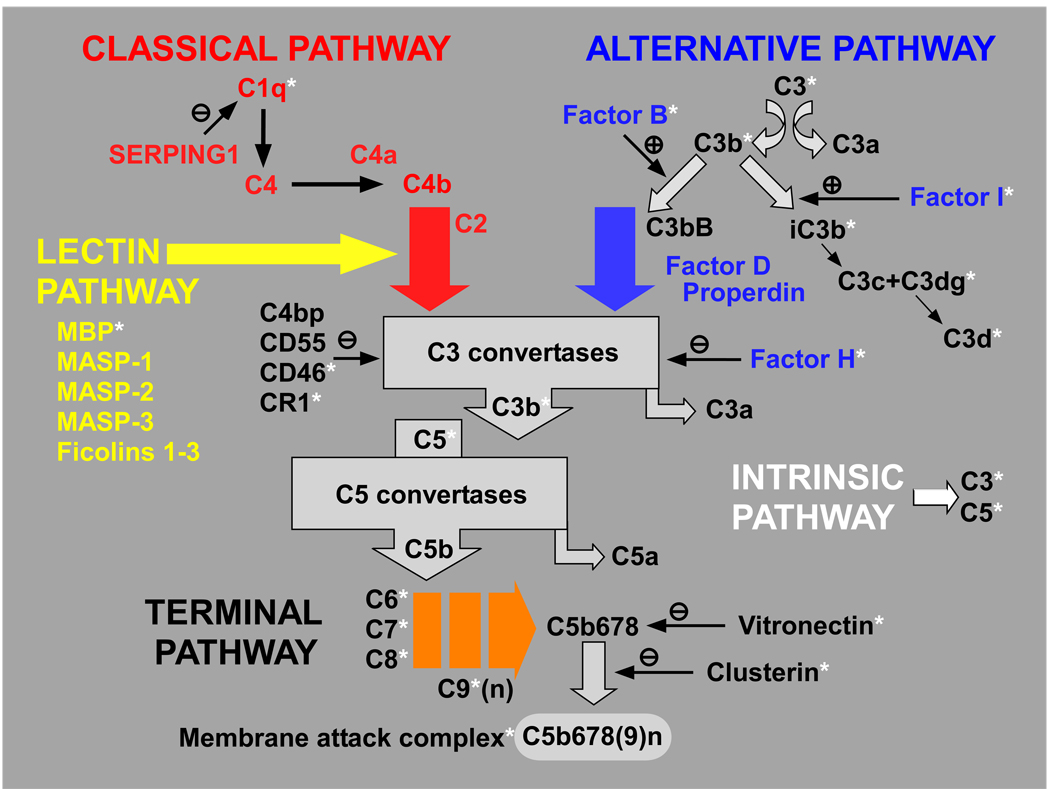
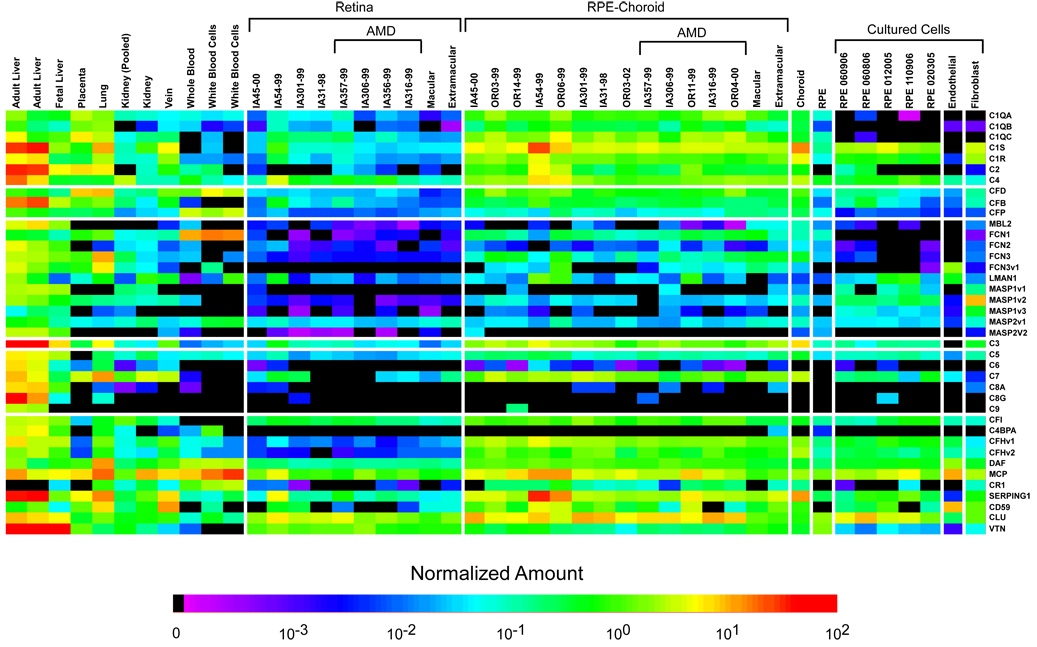
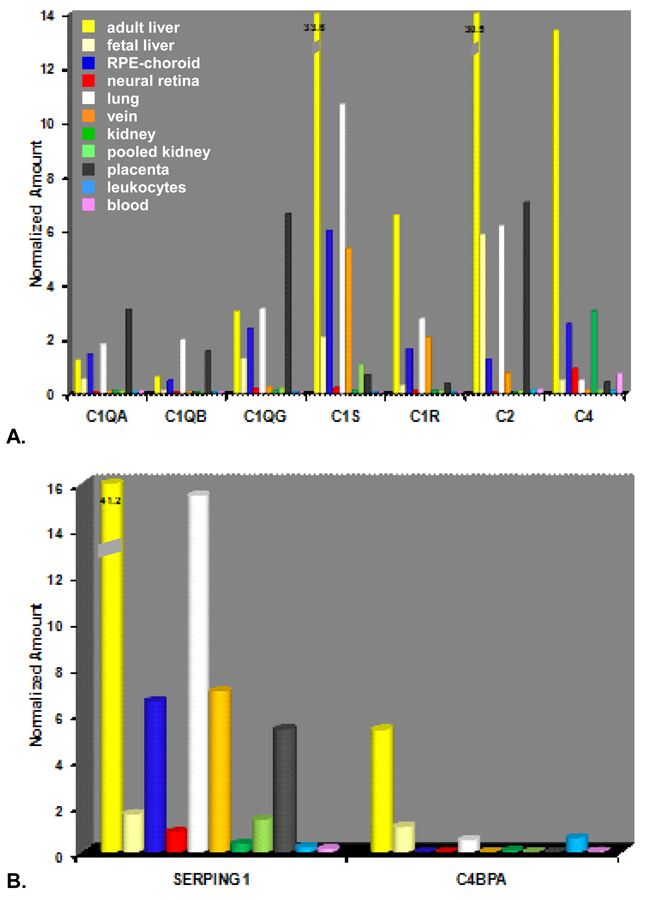
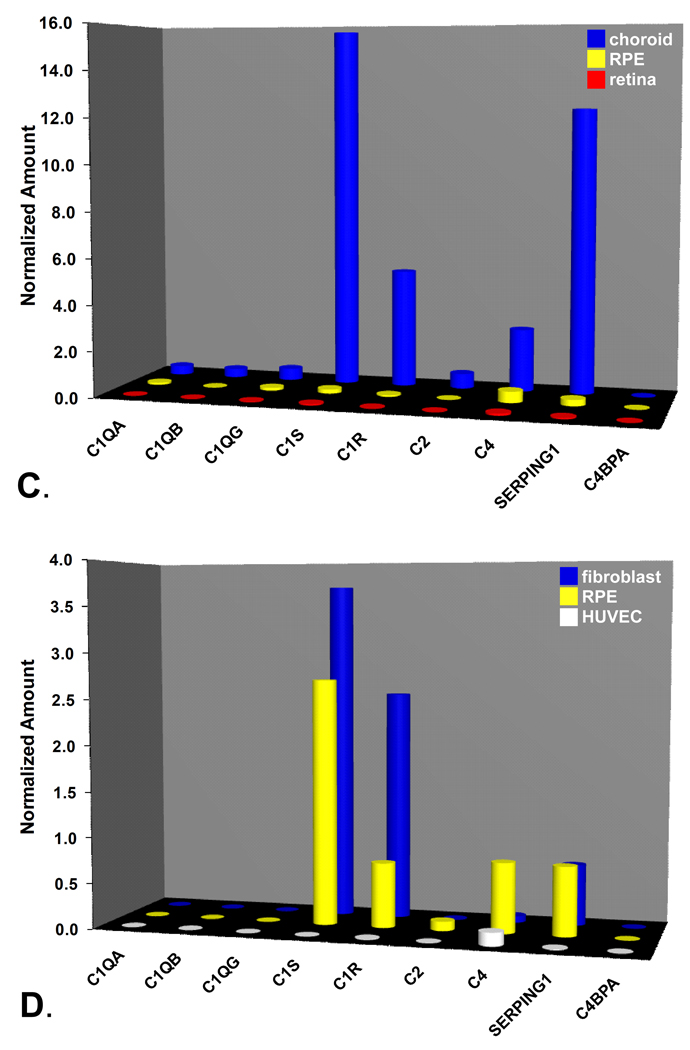
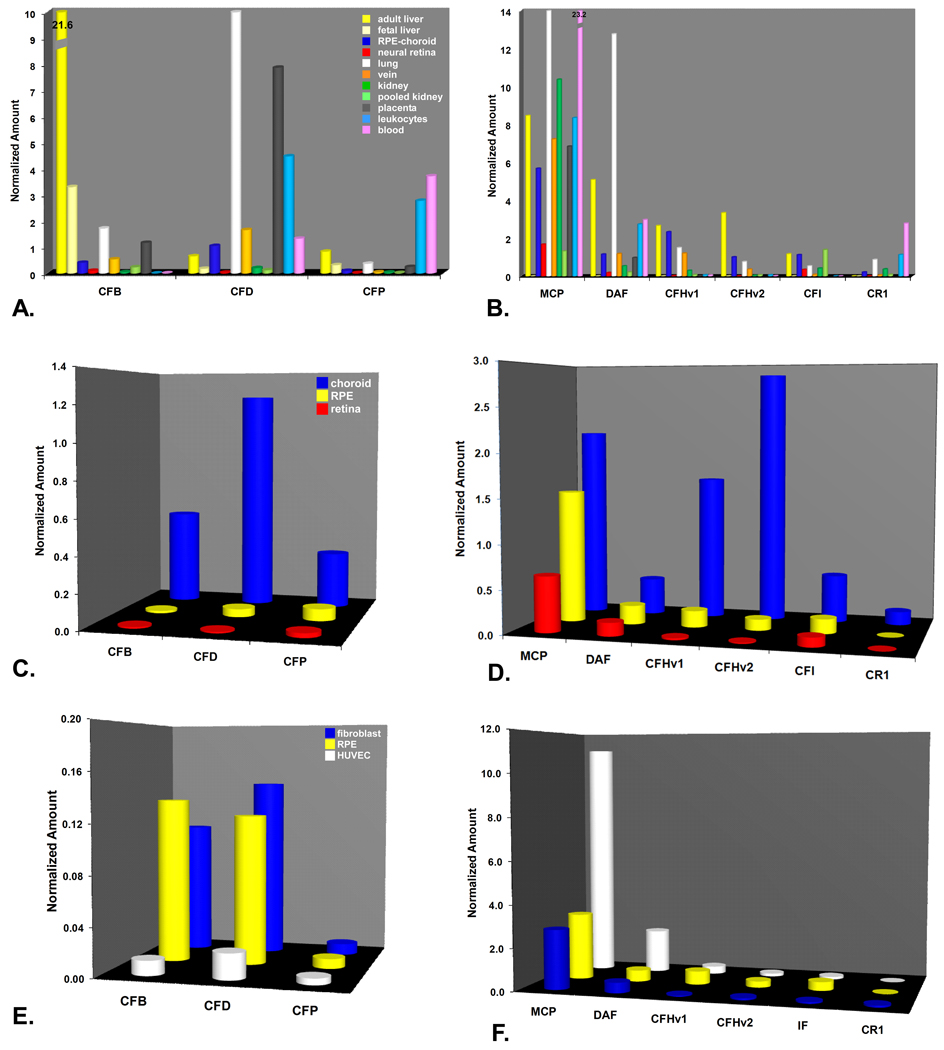
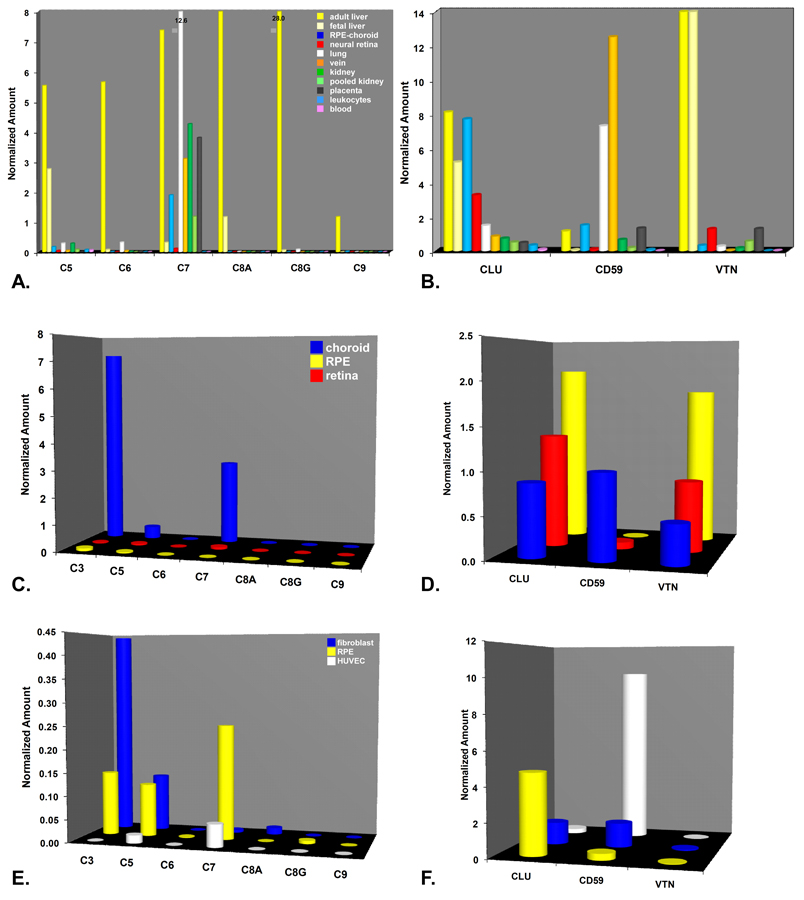
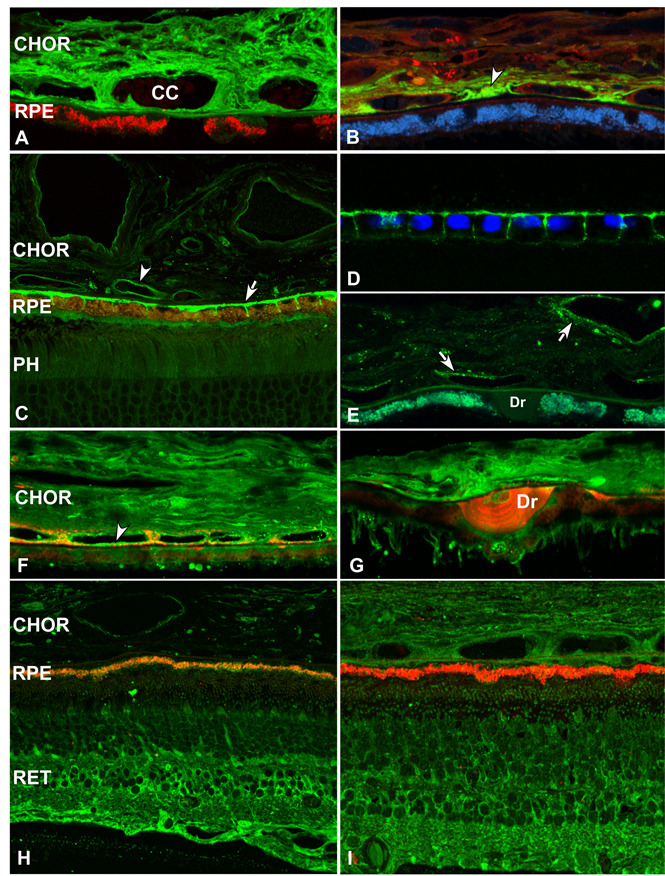
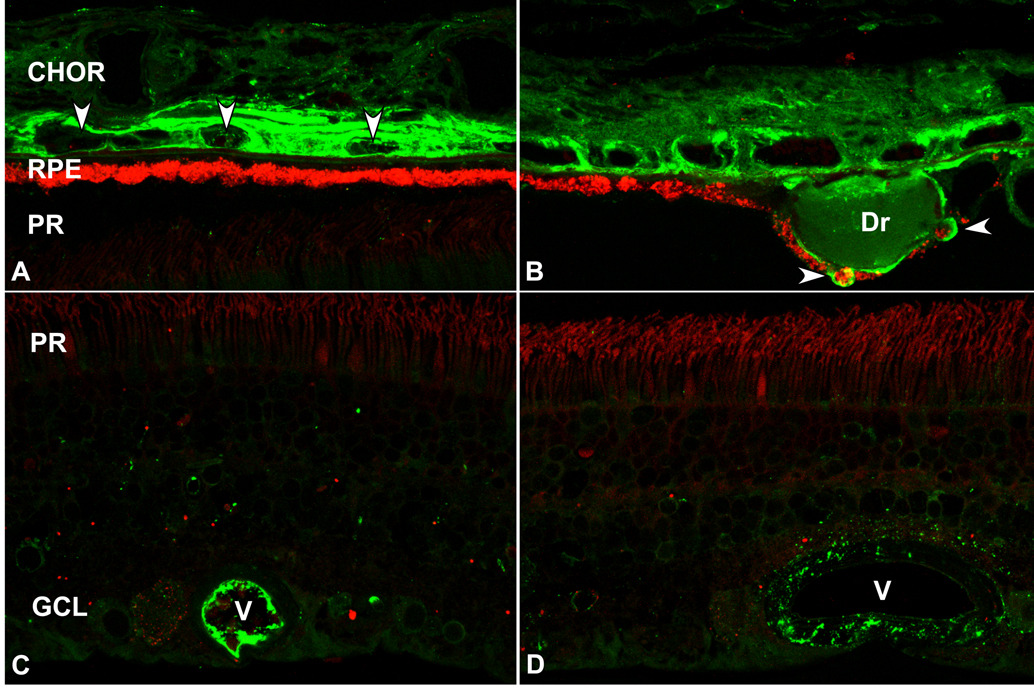
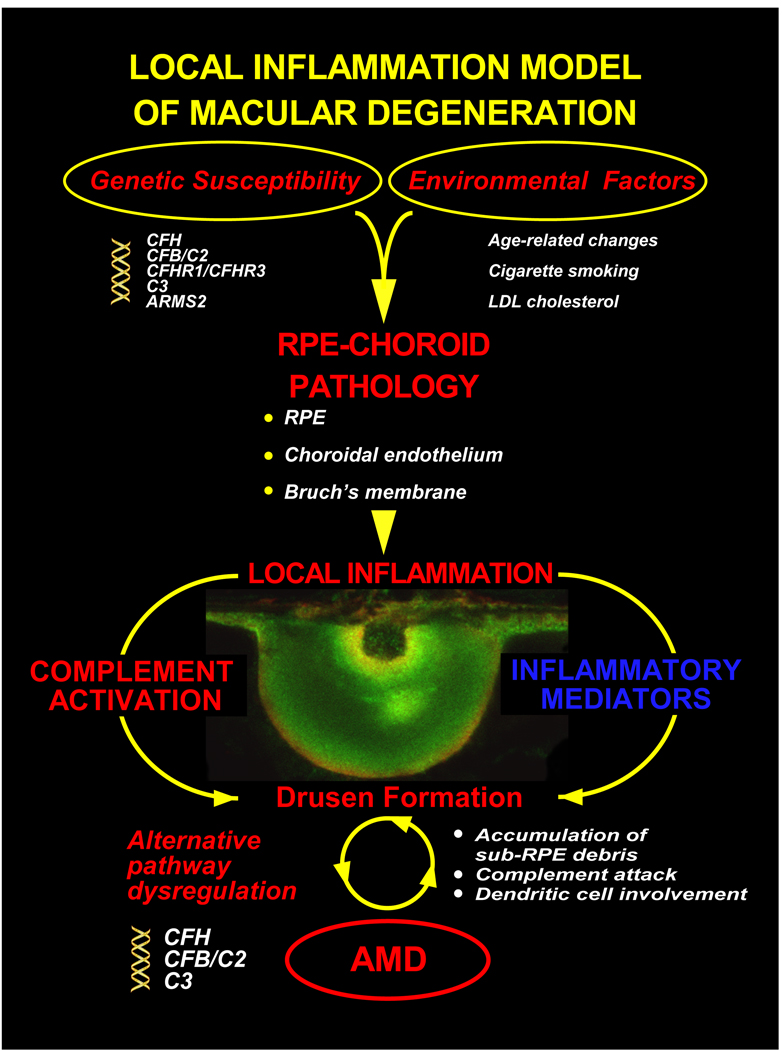
During the past ten years, dramatic advances have been made in unraveling the biological bases of age-related macular degeneration (AMD), the most common cause of irreversible blindness in western populations. In that timeframe, two distinct lines of evidence emerged which implicated chronic local inflammation and activation of the complement cascade in AMD pathogenesis. First, a number of complement system proteins, complement activators, and complement regulatory proteins were identified as molecular constituents of drusen, the hallmark extracellular deposits associated with early AMD. Subsequently, genetic studies revealed highly significant statistical associations between AMD and variants of several complement pathway-associated genes including: Complement factor H (CFH), complement factor H-related 1 and 3 (CFHR1 and CFHR3), complement factor B (CFB), complement component 2 (C2), and complement component 3 (C3). In this article, we revisit our original hypothesis that chronic local inflammatory and immune-mediated events at the level of Bruch's membrane play critical roles in drusen biogenesis and, by extension, in the pathobiology of AMD. Secondly, we report the results of a new screening for additional AMD-associated polymorphisms in a battery of 63 complement-related genes. Third, we identify and characterize the local complement system in the RPE-choroid complex - thus adding a new dimension of biological complexity to the role of the complement system in ocular aging and AMD. Finally, we evaluate the most salient, recent evidence that bears directly on the role of complement in AMD pathogenesis and progression. Collectively, these recent findings strongly re-affirm the importance of the complement system in AMD. They lay the groundwork for further studies that may lead to the identification of a transcriptional disease signature of AMD, and hasten the development of new therapeutic approaches that will restore the complement-modulating activity that appears to be compromised in genetically susceptible individuals.
Copyright 2009 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures
References
-
- Anderson DH, Mullins RF, Hageman GS, Johnson LV. A role for local inflammation in the formation of drusen in the aging eye. Am J Ophthalmol. 2002;134:411–431. - PubMed
-
- Anderson DH, Talaga KC, Rivest AJ, Barron E, Hageman GS, Johnson LV. Characterization of beta amyloid assemblies in drusen: the deposits associated with aging and age-related macular degeneration. Exp Eye Res. 2004;78:243–256. - PubMed
-
- Black S, Kushner I, Samols D. C-reactive Protein. J Biol Chem. 2004;279:48487–48490. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
