

How telomeres solve the end-protection problem
- PMID: 19965504
- PMCID: PMC2819049
- DOI: 10.1126/science.1170633
How telomeres solve the end-protection problem
Abstract
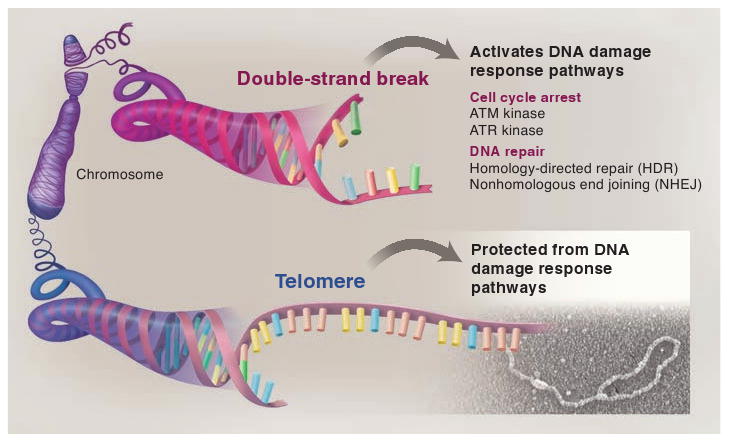
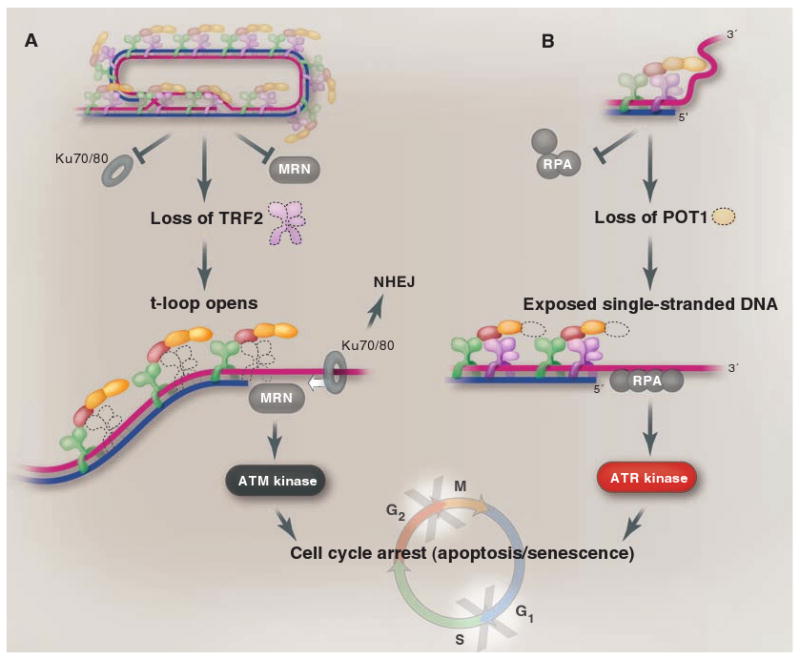
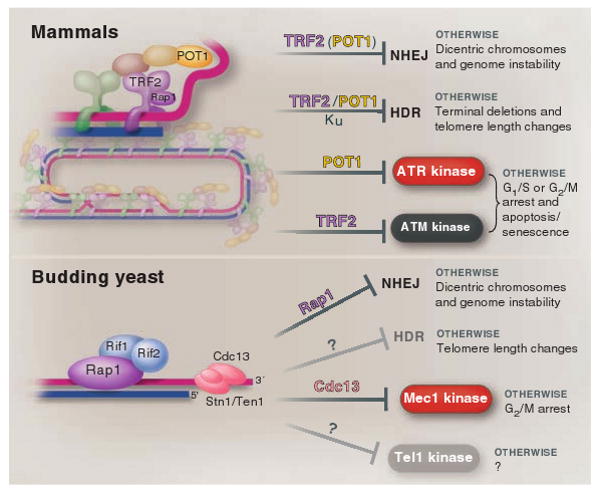
The ends of eukaryotic chromosomes have the potential to be mistaken for damaged or broken DNA and must therefore be protected from cellular DNA damage response pathways. Otherwise, cells might permanently arrest in the cell cycle, and attempts to "repair" the chromosome ends would have devastating consequences for genome integrity. This end-protection problem is solved by protein-DNA complexes called telomeres. Studies of mammalian cells have recently uncovered the mechanism by which telomeres disguise the chromosome ends. Comparison to unicellular eukaryotes reveals key differences in the DNA damage response systems that inadvertently threaten chromosome ends. Telomeres appear to be tailored to these variations, explaining their variable structure and composition.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
