

The NDR kinase DBF-2 is involved in regulation of mitosis, conidial development, and glycogen metabolism in Neurospora crassa
- PMID: 19966031
- PMCID: PMC2863412
- DOI: 10.1128/EC.00230-09
The NDR kinase DBF-2 is involved in regulation of mitosis, conidial development, and glycogen metabolism in Neurospora crassa
Abstract
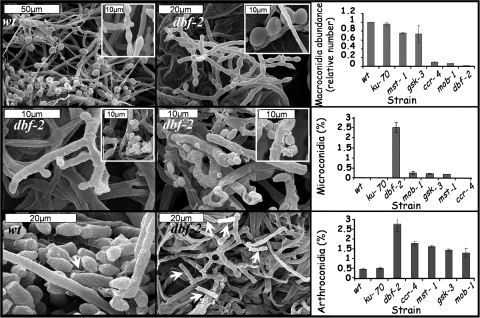
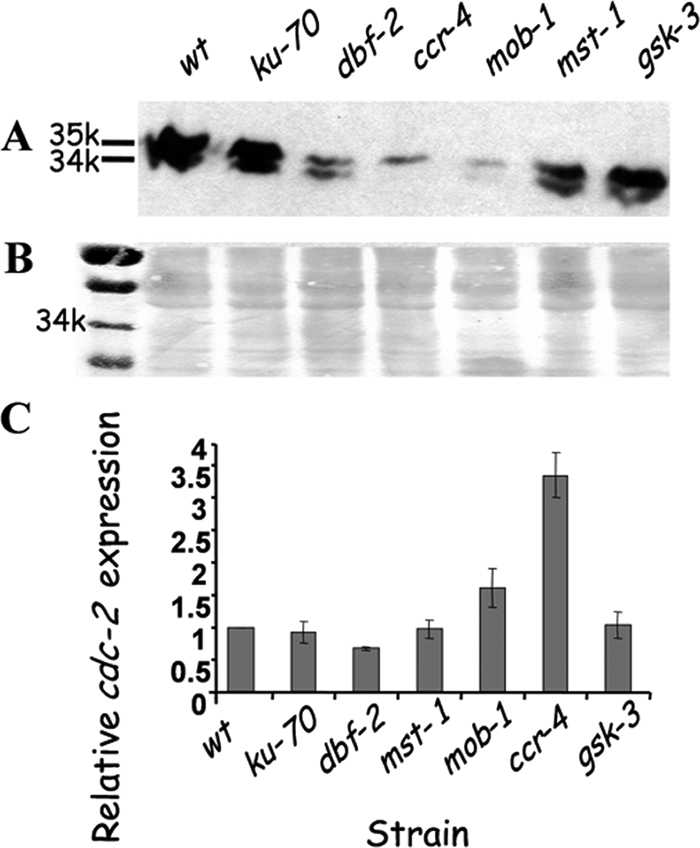
Neurospora crassa dbf-2 encodes an NDR (nuclear Dbf2-related) protein kinase, homologous to LATS1, a core component of the Hippo pathway. This pathway plays important roles in restraining cell proliferation and promoting apoptosis in differentiating cells. Here, we demonstrate that DBF-2 is involved in three fundamental processes in a filamentous fungus: cell cycle regulation, glycogen biosynthesis, and conidiation. DBF-2 is predominantly localized to the nucleus, and most (approximately 60%) dbf-2 null mutant nuclei are delayed in mitosis, indicating that DBF-2 activity is required for properly completing the cell cycle. The dbf-2 mutant exhibits reduced basal hyphal extension rates accompanied by a carbon/nitrogen ratio-dependent bursting of hyphal tips, vast glycogen leakage, defects in aerial hypha formation, and impairment of all three asexual conidiation pathways in N. crassa. Our findings also indicate that DBF-2 is essential for sexual reproduction in a filamentous fungus. Defects in other Hippo and glycogen metabolism pathway components (mob-1, ccr-4, mst-1, and gsk-3) share similar phenotypes such as mitotic delay and decreased CDC-2 (cell division cycle 2) protein levels, massive hyphal swellings, hyphal tip bursting, glycogen leakage, and impaired conidiation. We propose that DBF-2 functions as a link between Hippo and glycogen metabolism pathways.
Figures







References
-
- Borkovich K. A., Alex L. A., Yarden O., Freitag M., Turner G. E., Read N. D., Seiler S., Bell-Pedersen D., Paietta J., Plesofsky N., Plamann M., Goodrich-Tanrikulu M., Schulte U., Mannhaupt G., Nargang F. E., Radford A., Selitrennikoff C., Galagan J. E., Dunlap J. C., Loros J. J., Catcheside D., Inoue H., Aramayo R., Polymenis M., Selker E. U., Sachs M. S., Marzluf G. A., Paulsen I., Davis R., Ebbole D. J., Zelter A., Kalkman E. R., O'Rourke R., Bowring F., Yeadon J., Ishii C., Suzuki K., Sakai W., Pratt R. 2004. Lessons from the genome sequence of Neurospora crassa: tracing the path from genomic blueprint to multicellular organism. Microbiol. Mol. Biol. Rev. 68:1–108 - PMC - PubMed
-
- Bothos J., Tuttle R. L., Ottey M., Luca F. C., Halazonetis T. D. 2005. Human LATS1 is a mitotic exit network kinase. Cancer Res. 65:6568–6575 - PubMed
-
- Bradford M. M. 1976. A rapid and sensitive method for the quantitation of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248–254 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
