

Sleep-wake regulation and hypocretin-melatonin interaction in zebrafish
- PMID: 19966231
- PMCID: PMC2799794
- DOI: 10.1073/pnas.906637106
Sleep-wake regulation and hypocretin-melatonin interaction in zebrafish
Abstract
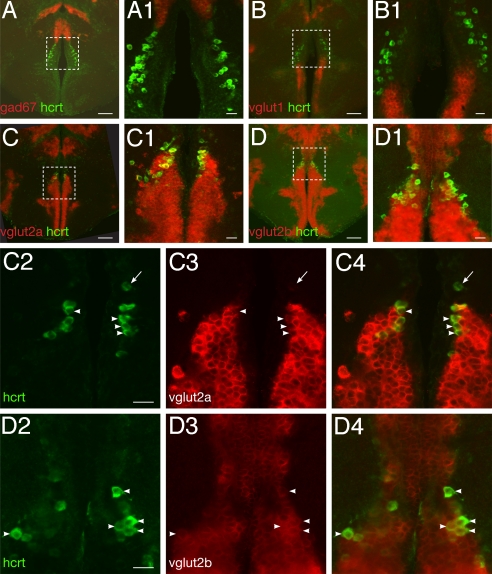
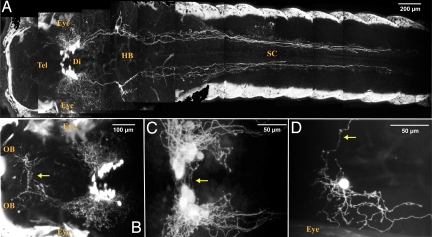
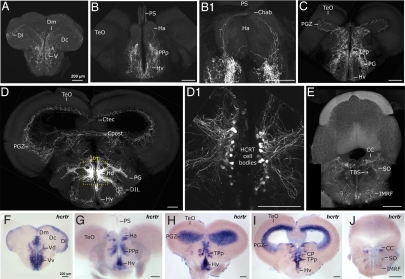
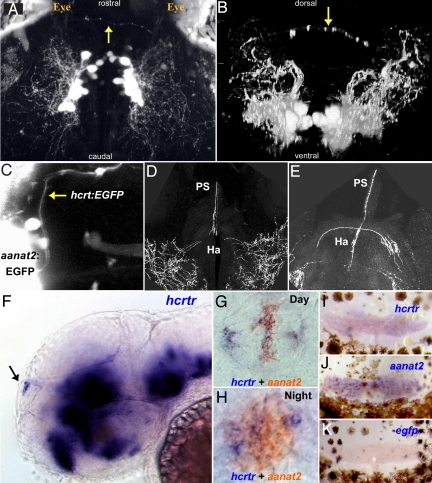
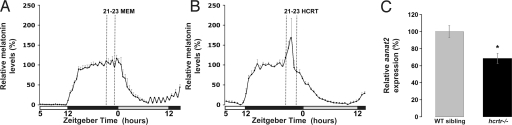
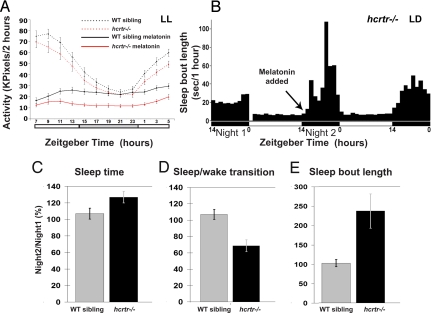
In mammals, hypocretin/orexin (HCRT) neuropeptides are important sleep-wake regulators and HCRT deficiency causes narcolepsy. In addition to fragmented wakefulness, narcoleptic mammals also display sleep fragmentation, a less understood phenotype recapitulated in the zebrafish HCRT receptor mutant (hcrtr-/-). We therefore used zebrafish to study the potential mediators of HCRT-mediated sleep consolidation. Similar to mammals, zebrafish HCRT neurons express vesicular glutamate transporters indicating conservation of the excitatory phenotype. Visualization of the entire HCRT circuit in zebrafish stably expressing hcrt:EGFP revealed parallels with established mammalian HCRT neuroanatomy, including projections to the pineal gland, where hcrtr mRNA is expressed. As pineal-produced melatonin is a major sleep-inducing hormone in zebrafish, we further studied how the HCRT and melatonin systems interact functionally. mRNA level of arylalkylamine-N-acetyltransferase (AANAT2), a key enzyme of melatonin synthesis, is reduced in hcrtr-/- pineal gland during the night. Moreover, HCRT perfusion of cultured zebrafish pineal glands induces melatonin release. Together these data indicate that HCRT can modulate melatonin production at night. Furthermore, hcrtr-/- fish are hypersensitive to melatonin, but not other hypnotic compounds. Subthreshold doses of melatonin increased the amount of sleep and consolidated sleep in hcrtr-/- fish, but not in the wild-type siblings. These results demonstrate the existence of a functional HCRT neurons-pineal gland circuit able to modulate melatonin production and sleep consolidation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
