

Collective dynamics in human and monkey sensorimotor cortex: predicting single neuron spikes
- PMID: 19966837
- PMCID: PMC2820252
- DOI: 10.1038/nn.2455
Collective dynamics in human and monkey sensorimotor cortex: predicting single neuron spikes
Abstract
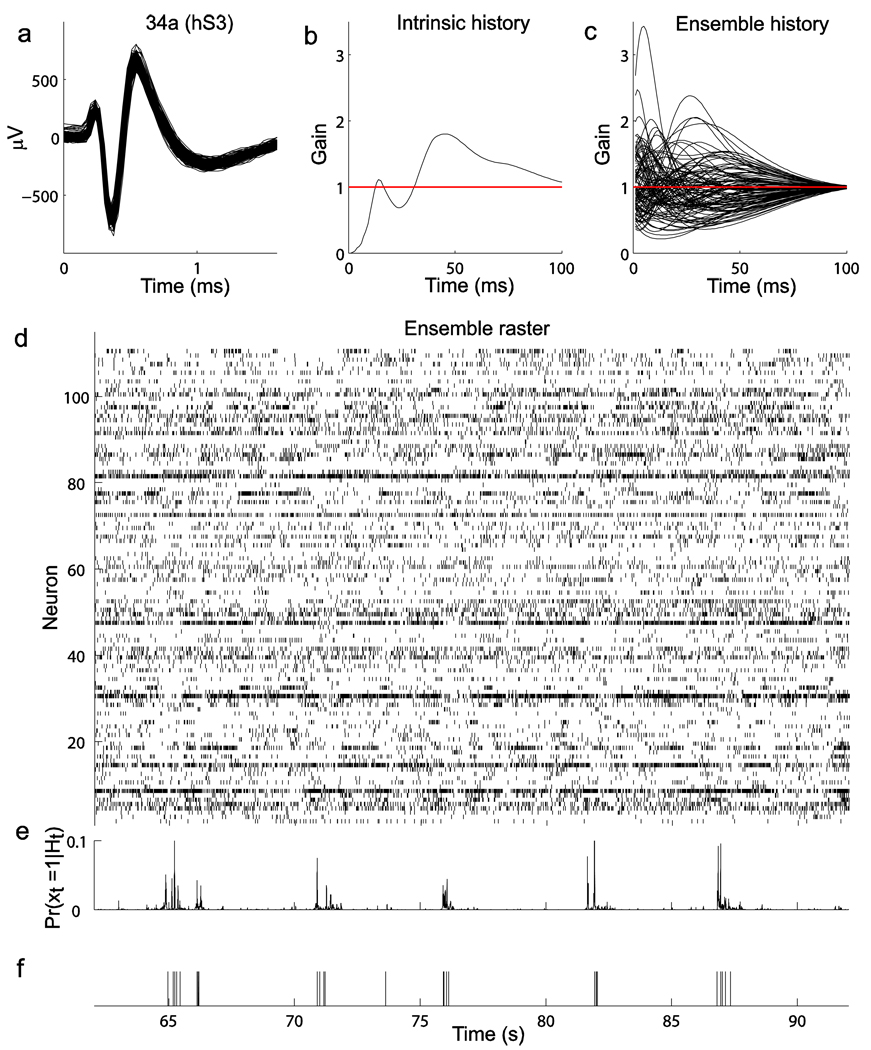
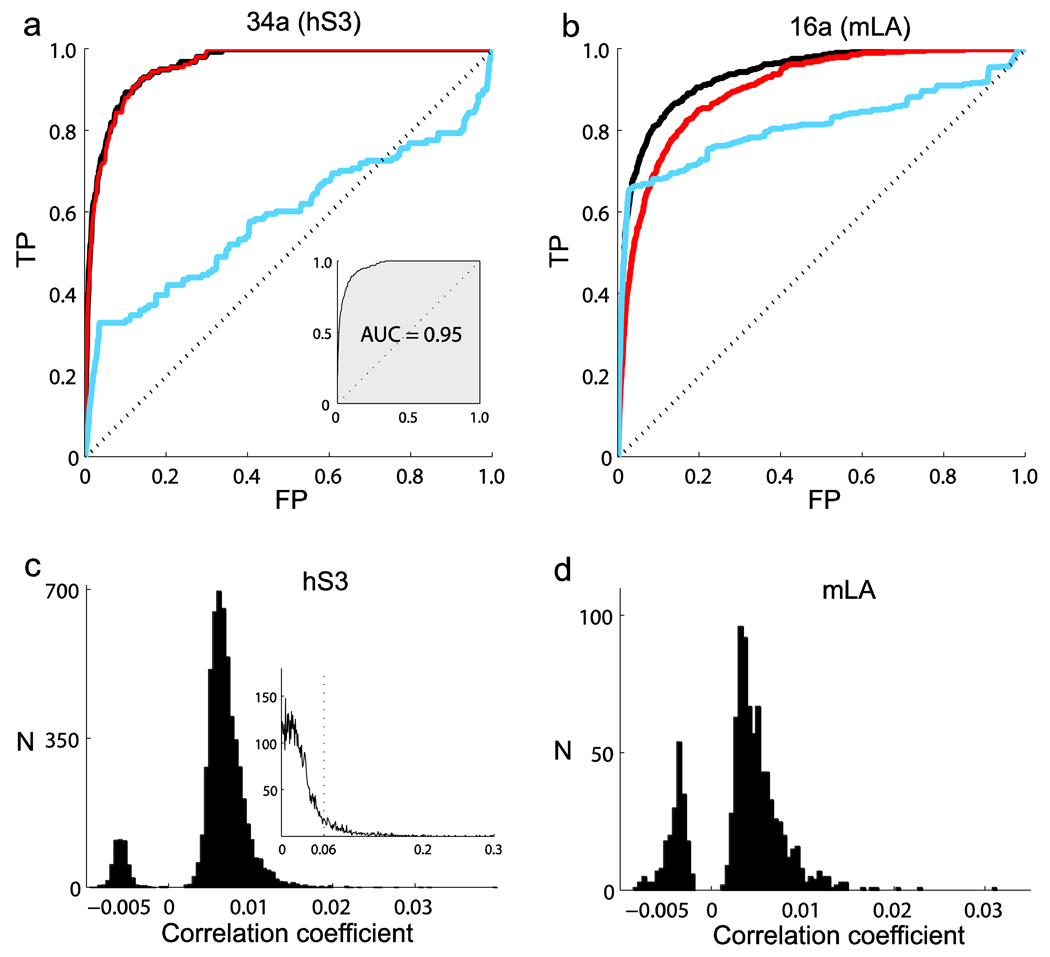
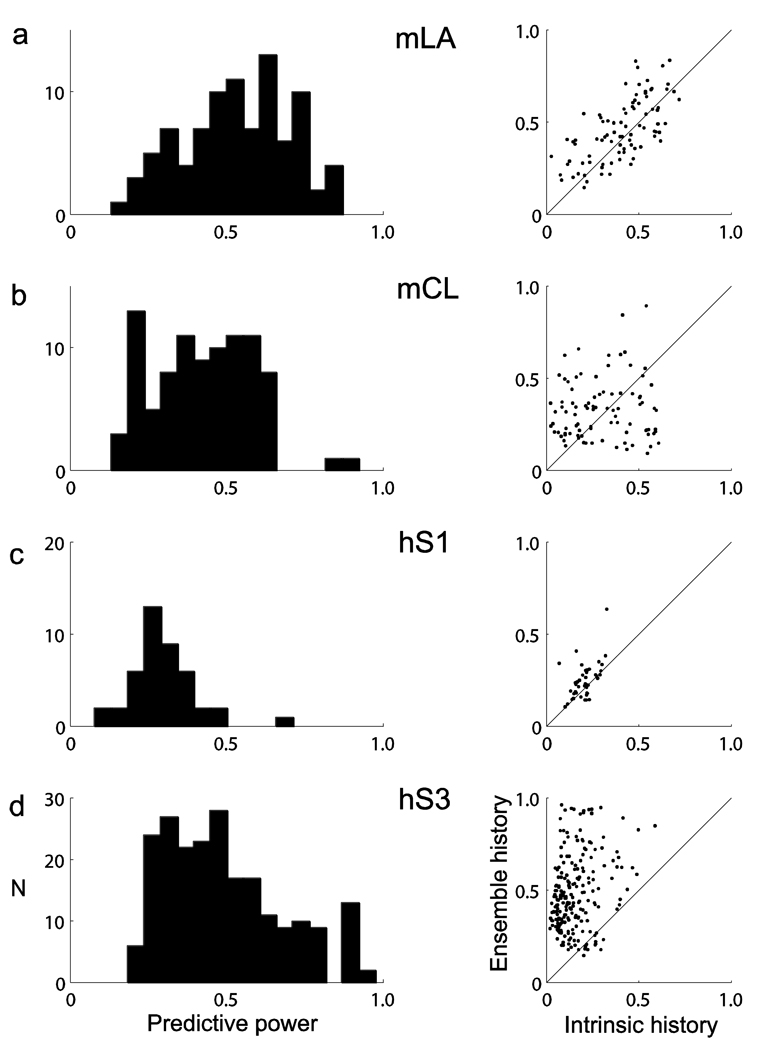
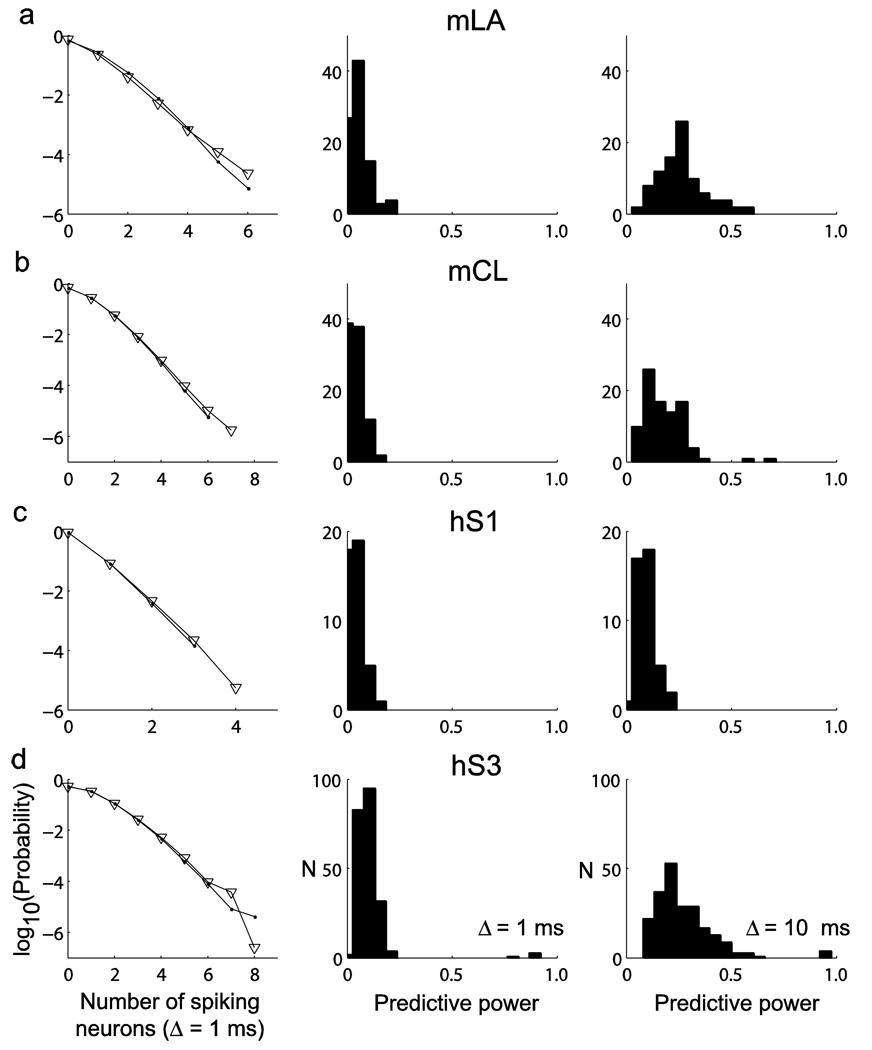
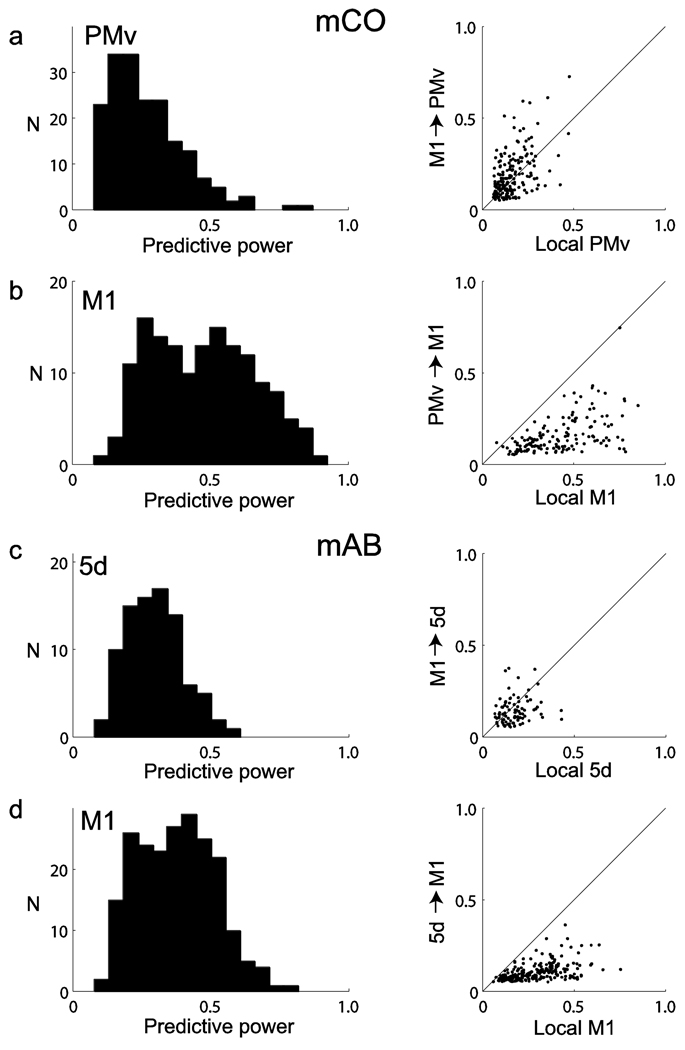
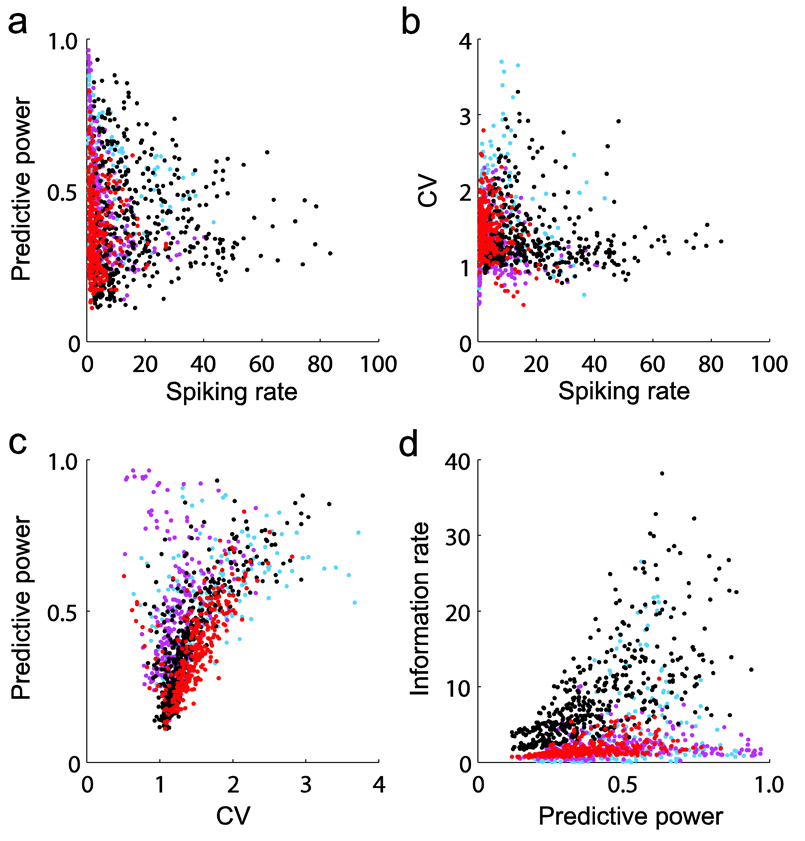
Coordinated spiking activity in neuronal ensembles, in local networks and across multiple cortical areas, is thought to provide the neural basis for cognition and adaptive behavior. Examining such collective dynamics at the level of single neuron spikes has remained, however, a considerable challenge. We found that the spiking history of small and randomly sampled ensembles (approximately 20-200 neurons) could predict subsequent single neuron spiking with substantial accuracy in the sensorimotor cortex of humans and nonhuman behaving primates. Furthermore, spiking was better predicted by the ensemble's history than by the ensemble's instantaneous state (Ising models), emphasizing the role of temporal dynamics leading to spiking. Notably, spiking could be predicted not only by local ensemble spiking histories, but also by spiking histories in different cortical areas. These strong collective dynamics may provide a basis for understanding cognition and adaptive behavior at the level of coordinated spiking in cortical networks.
Figures
References
-
- Braitenberg V, Schüz A. Cortex: Statistics and Geometry of Neuronal Connectivity. New York: Springer-Verlag; 1998.
-
- Elston GN, Rockland KS. The pyramidal cell of the sensorimotor cortex of the macaque monkey: phenotypic variation. Cereb. Cortex. 2002;12:1071–1078. - PubMed
-
- Dayan P, Abbott LF. Theoretical Neuroscience. Cambridge, Massachusetts: MIT Press; 2001.
-
- Destexhe A, Rudolph M, Paré D. The high-conductance state of neocortical neurons in vivo. Nat. Rev. Neurosci. 2003;4:739–751. - PubMed
-
- Arieli A, Sterkin A, Grinvald A, Aertsen A. Dynamics of ongoing activity: explanation of the large variability in evoked cortical responses. Science. 1996;273:1868–1871. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
