

Spike integration and cellular memory in a rhythmic network from Na+/K+ pump current dynamics
- PMID: 19966842
- PMCID: PMC2839136
- DOI: 10.1038/nn.2444
Spike integration and cellular memory in a rhythmic network from Na+/K+ pump current dynamics
Abstract
The output of a neural circuit results from an interaction between the intrinsic properties of neurons in the circuit and the features of the synaptic connections between them. The plasticity of intrinsic properties has been primarily attributed to modification of ion channel function and/or number. We have found a mechanism for intrinsic plasticity in rhythmically active Drosophila neurons that was not based on changes in ion conductance. Larval motor neurons had a long-lasting, sodium-dependent afterhyperpolarization (AHP) following bursts of action potentials that was mediated by the electrogenic activity of Na(+)/K(+) ATPase. This AHP persisted for multiple seconds following volleys of action potentials and was able to function as a pattern-insensitive integrator of spike number that was independent of external calcium. This current also interacted with endogenous Shal K(+) conductances to modulate spike timing for multiple seconds following rhythmic activity, providing a cellular memory of network activity on a behaviorally relevant timescale.
Figures
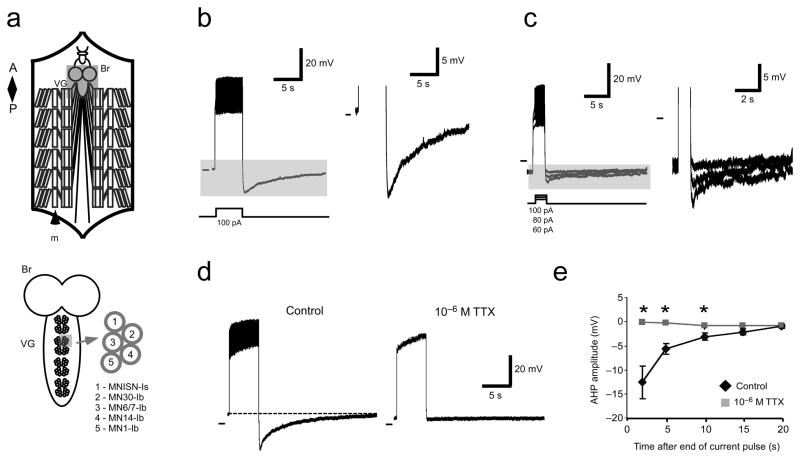
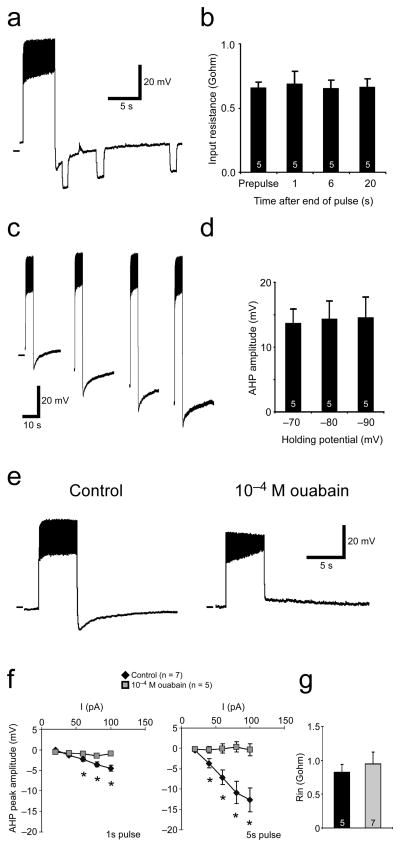
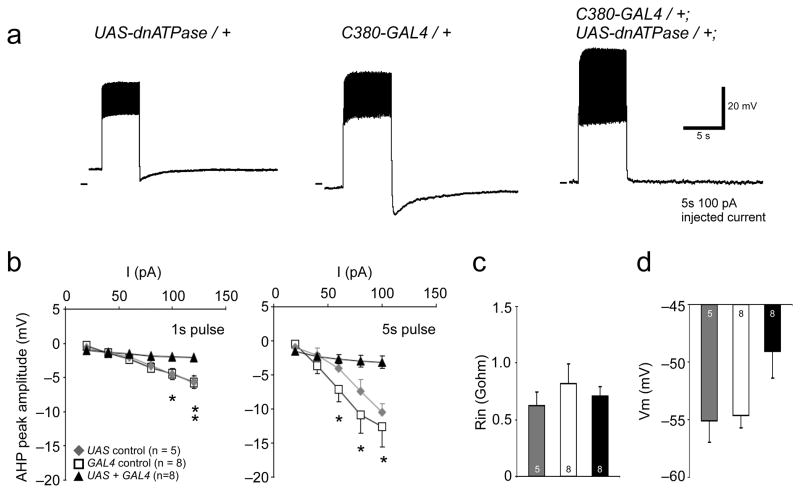
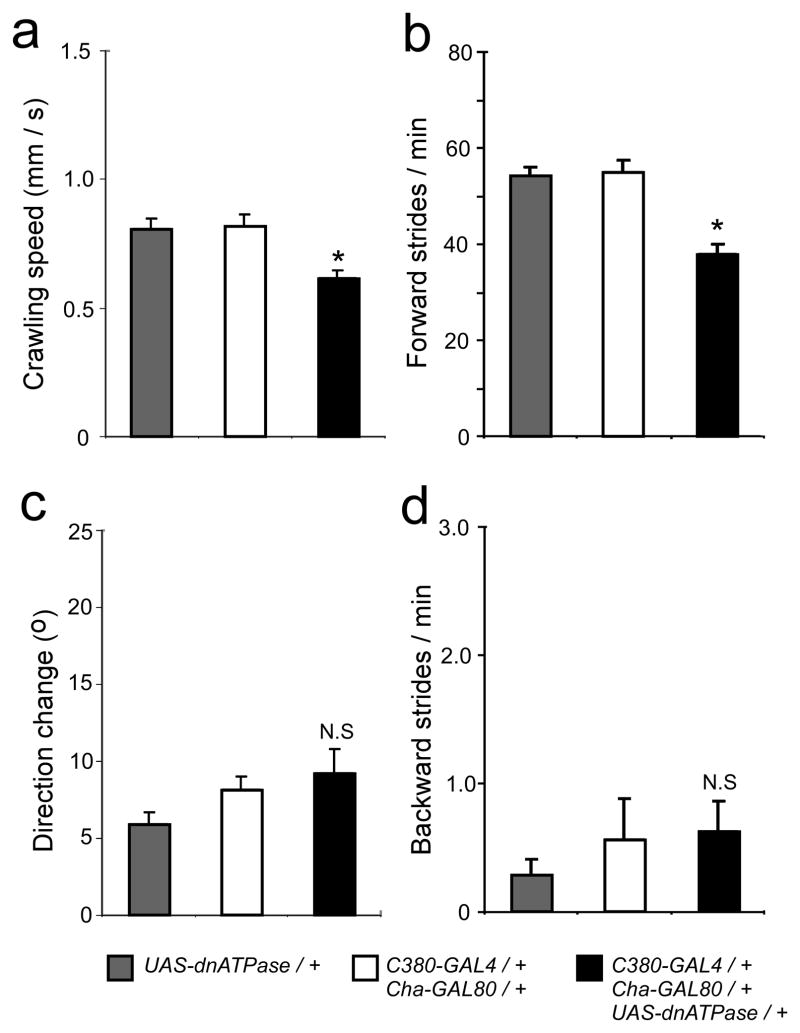


Comment in
-
Ion pumps get more glamorous.Nat Neurosci. 2010 Jan;13(1):4-5. doi: 10.1038/nn0110-4. Nat Neurosci. 2010. PMID: 20033078 No abstract available.
References
-
- Cavazzini M, Bliss T, Emptage N. Ca2+ and synaptic plasticity. Cell Calcium. 2005;38:355–367. - PubMed
-
- Cudmore RH, Turrigiano GG. Long-term potentiation of intrinsic excitability in LV visual cortical neurons. J Neurophysiol. 2004;92:341–348. - PubMed
-
- Turrigiano G, Abbott LF, Marder E. Activity-dependent changes in the intrinsic properties of cultured neurons. Science. 1994;264:974–977. - PubMed
-
- Koch C. Biophysics of Computation. Oxford University Press; Oxford: 1999.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
