

Asymmetric cross-regulation between the nitrate-responsive NarX-NarL and NarQ-NarP two-component regulatory systems from Escherichia coli K-12
- PMID: 19968795
- PMCID: PMC3034140
- DOI: 10.1111/j.1365-2958.2009.06987.x
Asymmetric cross-regulation between the nitrate-responsive NarX-NarL and NarQ-NarP two-component regulatory systems from Escherichia coli K-12
Abstract
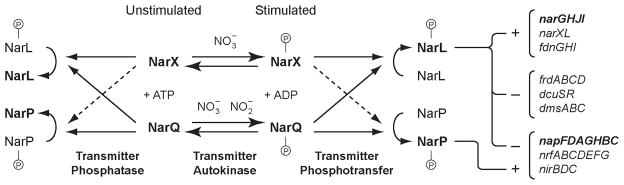
The NarX-NarL and NarQ-NarP sensor-response regulator pairs control Escherichia coli gene expression in response to nitrate and nitrite. Previous analysis suggests that the Nar two-component systems form a cross-regulation network in vivo. Here we report on the kinetics of phosphoryl transfer between different sensor-regulator combinations in vitro. NarX exhibited a noticeable kinetic preference for NarL over NarP, whereas NarQ exhibited a relatively slight kinetic preference for NarL. These findings were substantiated in reactions containing one sensor and both response regulators, or with two sensors and a single response regulator. We isolated 21 NarX mutants with missense substitutions in the cytoplasmic central and transmitter modules. These confer phenotypes that reflect defects in phospho-NarL dephosphorylation. Five of these mutants, all with substitutions in the transmitter DHp domain, also exhibited NarP-blind phenotypes. Phosphoryl transfer assays in vitro confirmed that these NarX mutants have defects in catalysing NarP phosphorylation. By contrast, the corresponding NarQ mutants conferred phenotypes indicating comparable interactions with both NarP and NarL. Our overall results reveal asymmetry in the Nar cross-regulation network, such that NarQ interacts similarly with both response regulators, whereas NarX interacts preferentially with NarL.
Figures






References
-
- Alves R, Savageau MA. Comparative analysis of prototype two-component systems with either bifunctional or monofunctional sensors: differences in molecular structure and physiological function. Mol Microbiol. 2003;48:25–51. - PubMed
-
- Cai SJ, Inouye M. EnvZ-OmpR interaction and osmoregulation in Escherichia coli. J Biol Chem. 2002;277:24155–24161. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
