

Novel angiogenesis inhibitory activity in cinnamon extract blocks VEGFR2 kinase and downstream signaling
- PMID: 19969552
- PMCID: PMC3105590
- DOI: 10.1093/carcin/bgp292
Novel angiogenesis inhibitory activity in cinnamon extract blocks VEGFR2 kinase and downstream signaling
Abstract
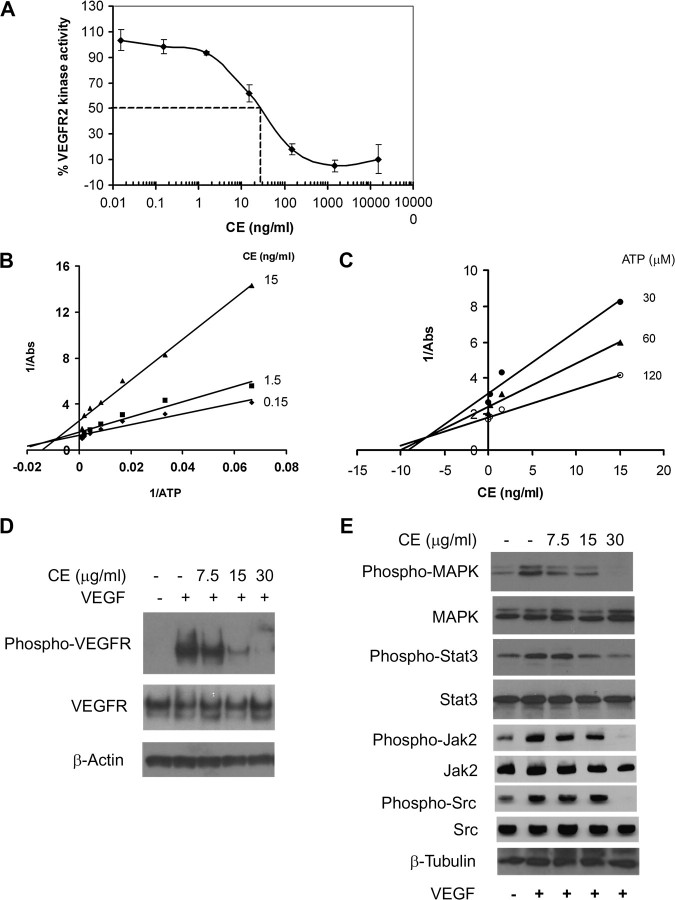
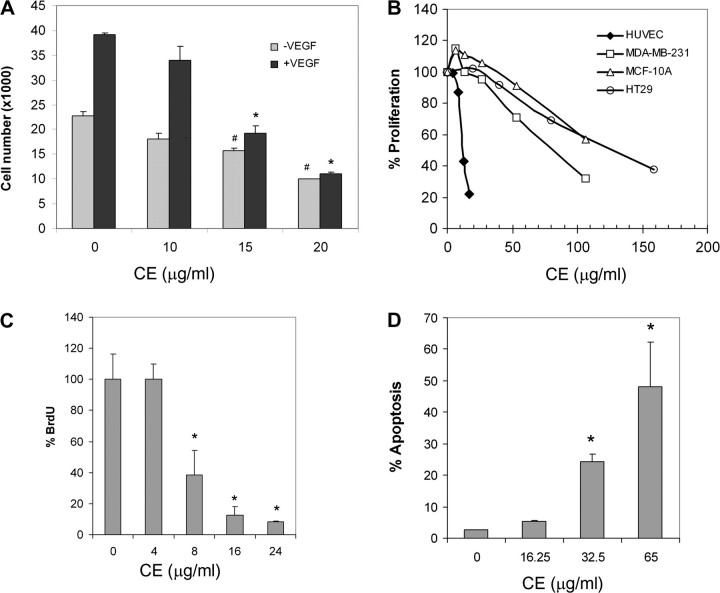
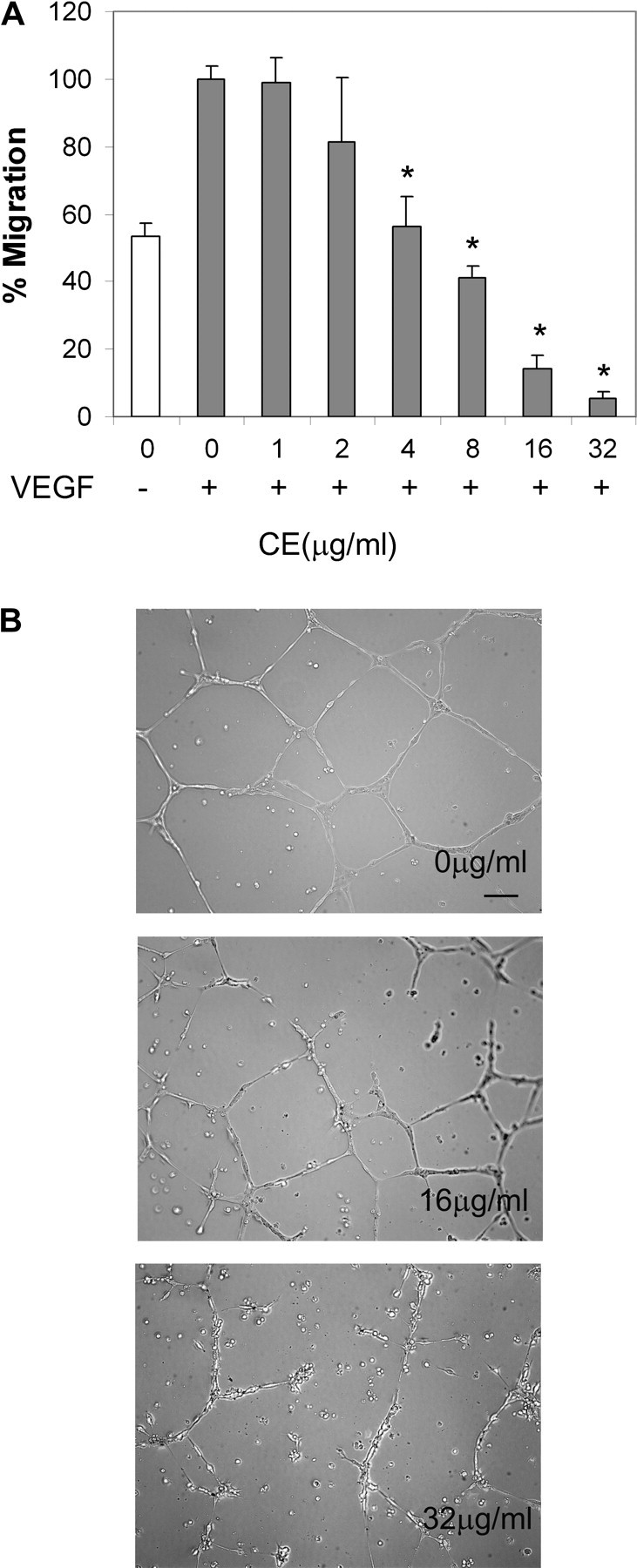
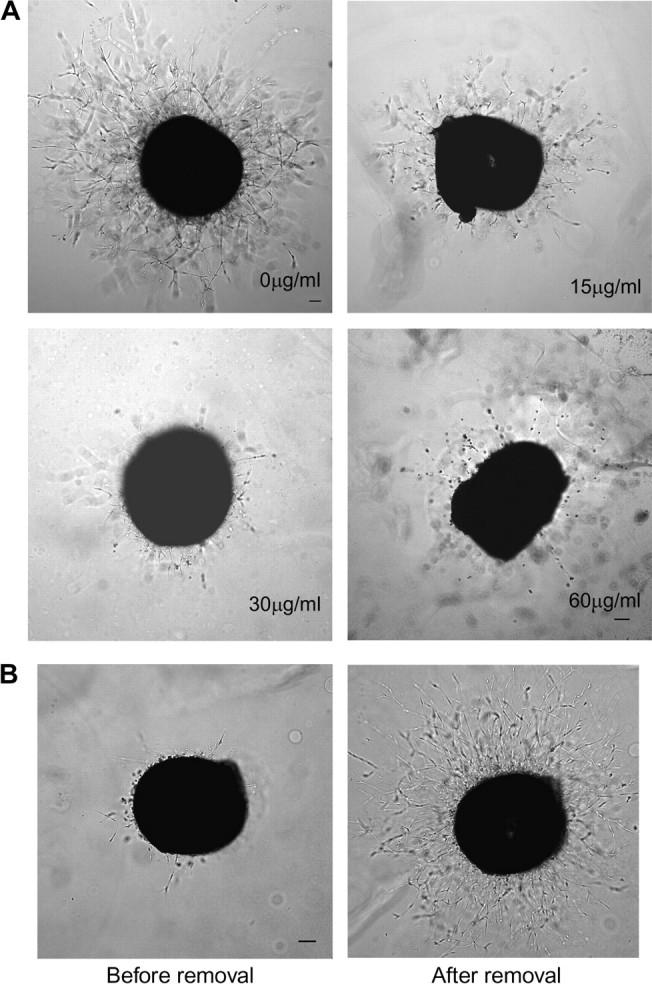
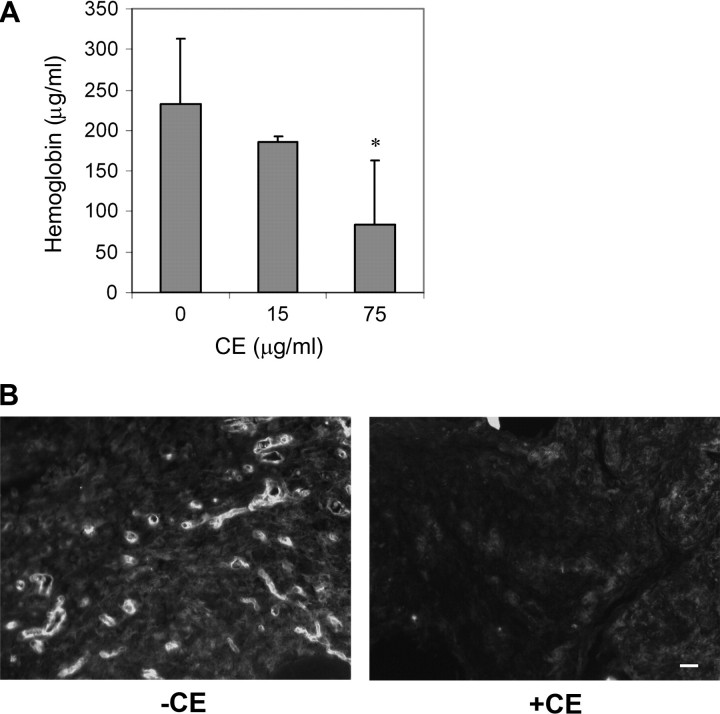
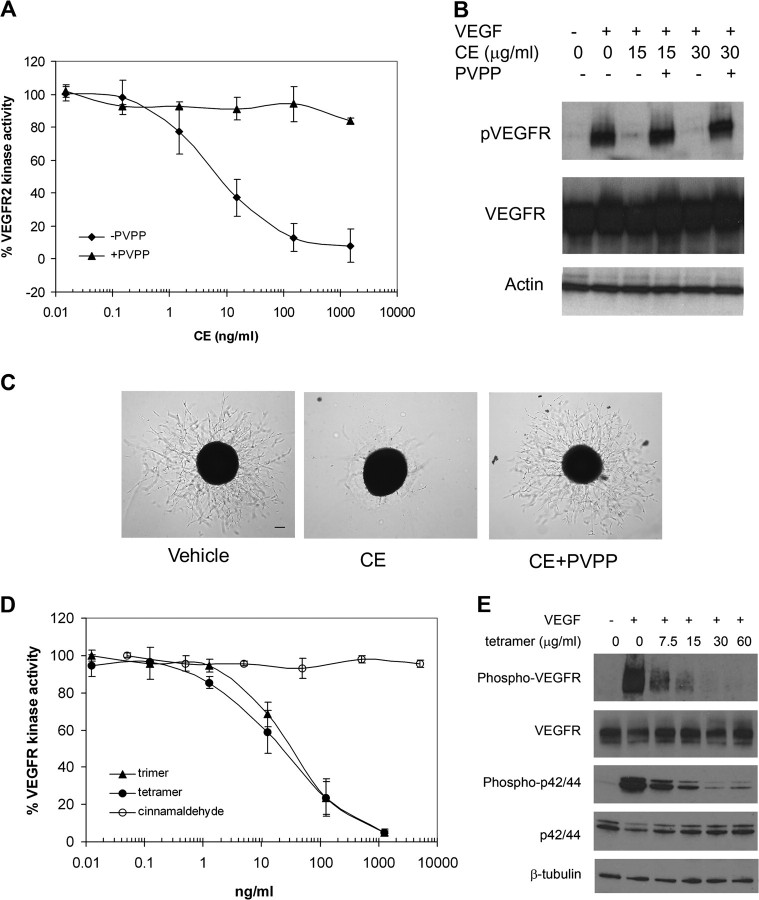
As a critical factor in the induction of angiogenesis, vascular endothelial growth factor (VEGF) has become an attractive target for anti-angiogenesis treatment. However, the side effects associated with most anti-VEGF agents limit their chronic use. Identification of naturally occurring VEGF inhibitors derived from diet is a potential alternative approach, with the advantage of known safety. To isolate natural inhibitors of VEGF, we established an in vitro tyrosine kinase assay to screen for diet-based agents that suppress VEGFR2 kinase activity. We found that a water-based extract from cinnamon (cinnamon extract, CE), one of the oldest and most popular spices, was a potent inhibitor of VEGFR2 kinase activity, directly inhibiting kinase activity of purified VEGFR2 as well as mitogen-activated protein kinase- and Stat3-mediated signaling pathway in endothelial cells. As a result, CE inhibited VEGF-induced endothelial cell proliferation, migration and tube formation in vitro, sprout formation from aortic ring ex vivo and tumor-induced blood vessel formation in vivo. Depletion of polyphenol from CE with polyvinylpyrrolidone abolished its anti-angiogenesis activity. While cinnamaldehyde, a component responsible for CE aroma, had little effect on VEGFR2 kinase activity, high-performance liquid chromatography-purified components of CE, procyanidin type A trimer (molecular weight, 864) and a tetramer (molecular weight, 1152) were found to inhibit kinase activity of purified VEGFR2 and VEGFR2 signaling, implicating procyanidin oligomers as active components in CE that inhibit angiogenesis. Our data revealed a novel activity in cinnamon and identified a natural VEGF inhibitor that could potentially be useful in cancer prevention and/or treatment.
Figures
References
-
- Kerbel R, et al. Clinical translation of angiogenesis inhibitors. Nat. Rev. Cancer. 2002;2:727–739. - PubMed
-
- Hanahan D, et al. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell. 1996;86:353–364. - PubMed
-
- Ferrara N. VEGF and the quest for tumor angiogenesis factors. Nat. Rev. Cancer. 2002;2:795–803. - PubMed
-
- Olsson A-K, et al. VEGF receptor signalling—in control of vascular function. Nat. Rev. Mol. Cell. Biol. 2006;7:359–371. - PubMed
-
- Ferrara N, et al. Angiogenesis as a therapeutic target. Nature. 2005;438:967–974. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
