

Ascorbate metabolism and the developmental demand for tartaric and oxalic acids in ripening grape berries
- PMID: 19995454
- PMCID: PMC2797797
- DOI: 10.1186/1471-2229-9-145
Ascorbate metabolism and the developmental demand for tartaric and oxalic acids in ripening grape berries
Abstract
Background: Fresh fruits are well accepted as a good source of the dietary antioxidant ascorbic acid (Asc, Vitamin C). However, fruits such as grapes do not accumulate exceptionally high quantities of Asc. Grapes, unlike most other cultivated fruits do however use Asc as a precursor for the synthesis of both oxalic (OA) and tartaric acids (TA). TA is a commercially important product in the wine industry and due to its acidifying effect on crushed juice it can influence the organoleptic properties of the wine. Despite the interest in Asc accumulation in fruits, little is known about the mechanisms whereby Asc concentration is regulated. The purpose of this study was to gain insights into Asc metabolism in wine grapes (Vitis vinifera c.v. Shiraz.) and thus ascertain whether the developmental demand for TA and OA synthesis influences Asc accumulation in the berry.
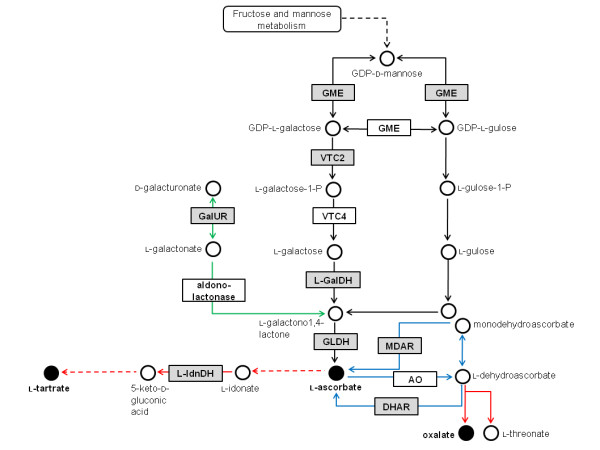
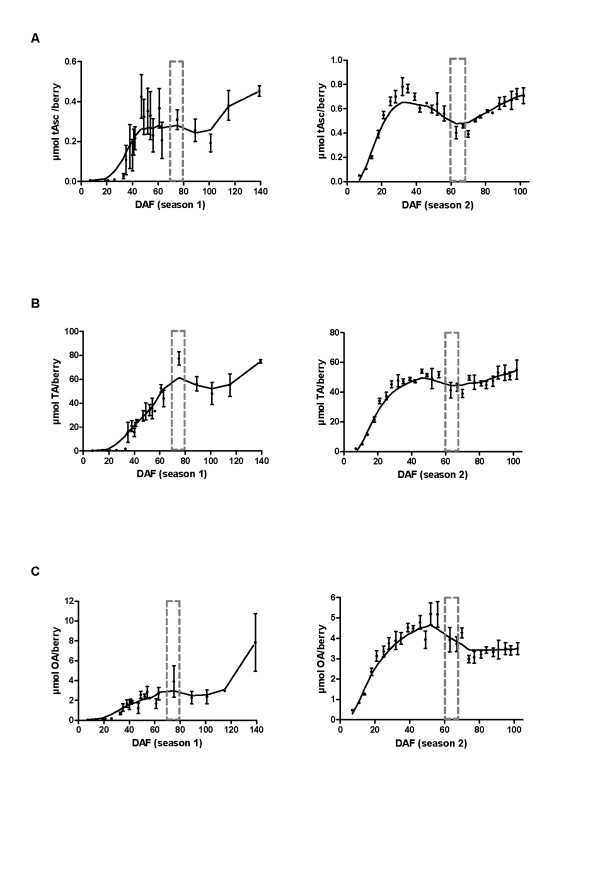
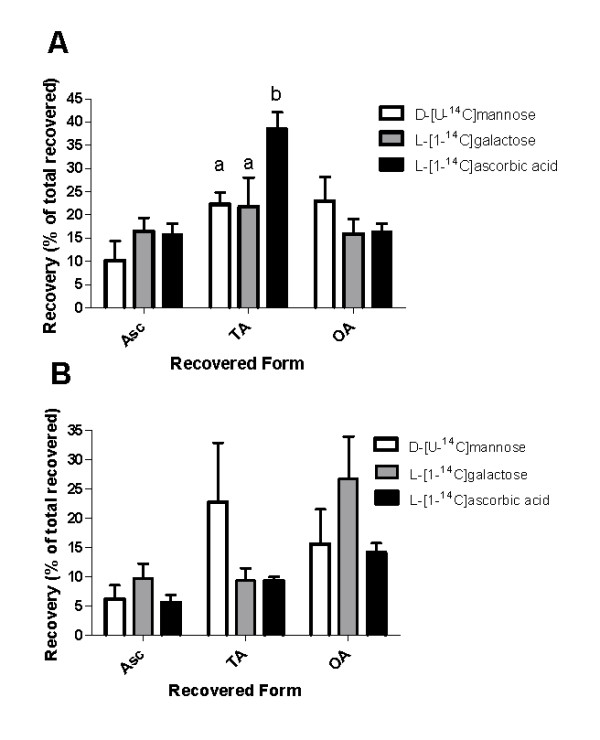
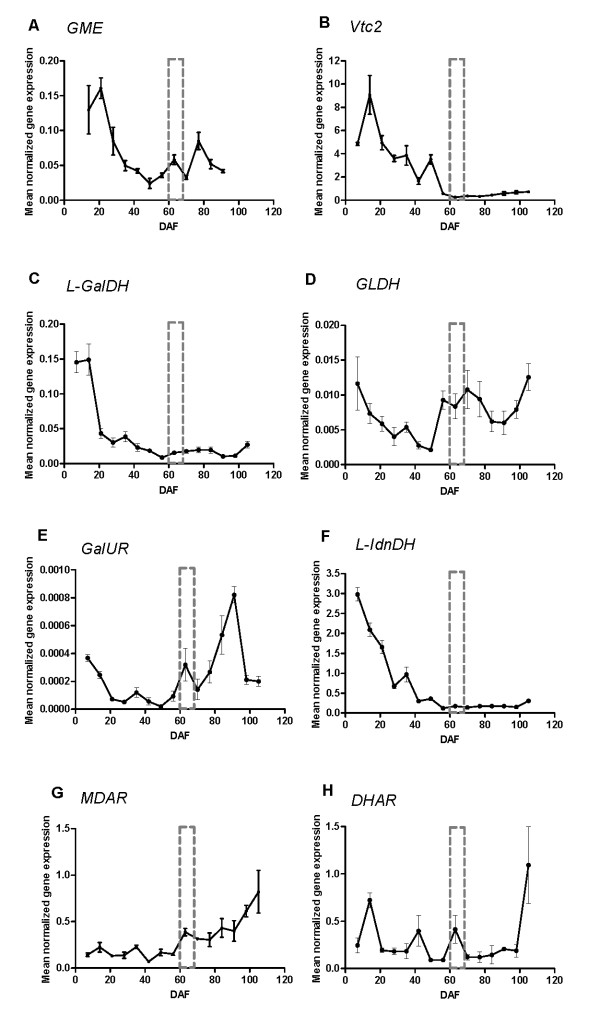
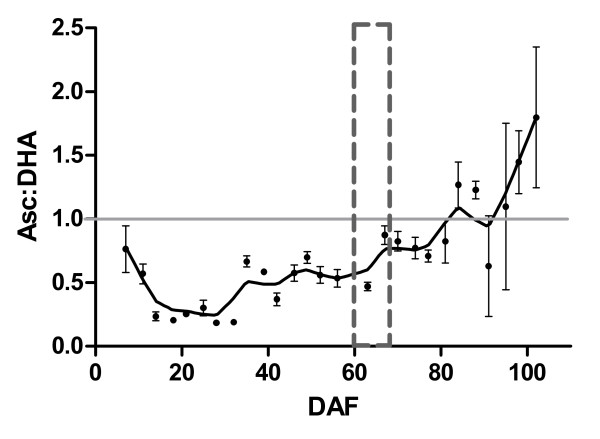
Results: We provide evidence for developmentally differentiated up-regulation of Asc biosynthetic pathways and subsequent fluctuations in Asc, TA and OA accumulation. Rapid accumulation of Asc and a low Asc to dehydroascorbate (DHA) ratio in young berries was co-ordinated with up-regulation of three of the primary Asc biosynthetic (Smirnoff-Wheeler) pathway genes. Immature berries synthesised Asc in-situ from the primary pathway precursors D-mannose and L-galactose. Immature berries also accumulated TA in early berry development in co-ordination with up-regulation of a TA biosynthetic gene. In contrast, ripe berries have up-regulated expression of the alternative Asc biosynthetic pathway gene D-galacturonic acid reductase with only residual expression of Smirnoff-Wheeler Asc biosynthetic pathway genes and of the TA biosynthetic gene. The ripening phase was further associated with up-regulation of Asc recycling genes, a secondary phase of increased accumulation of Asc and an increase in the Asc to DHA ratio.
Conclusion: We demonstrate strong developmental regulation of Asc biosynthetic, recycling and catabolic genes in grape berries. Integration of the transcript, radiotracer and metabolite data demonstrates that Asc and TA metabolism are developmentally regulated in grapevines; resulting in low accumulated levels of the biosynthetic intermediate Asc, and high accumulated levels of the metabolic end-product TA.
Figures
Similar articles
-
The role of light in the regulation of ascorbate metabolism during berry development in the cultivated grapevine Vitis vinifera L.J Sci Food Agric. 2011 Jul;91(9):1712-21. doi: 10.1002/jsfa.4376. Epub 2011 Mar 29. J Sci Food Agric. 2011. PMID: 21656772
-
Transcriptome analysis at four developmental stages of grape berry (Vitis vinifera cv. Shiraz) provides insights into regulated and coordinated gene expression.BMC Genomics. 2012 Dec 11;13:691. doi: 10.1186/1471-2164-13-691. BMC Genomics. 2012. PMID: 23227855 Free PMC article.
-
Berry skin development in Norton grape: distinct patterns of transcriptional regulation and flavonoid biosynthesis.BMC Plant Biol. 2011 Jan 10;11:7. doi: 10.1186/1471-2229-11-7. BMC Plant Biol. 2011. PMID: 21219654 Free PMC article.
-
Ascorbate as a biosynthetic precursor in plants.Ann Bot. 2007 Jan;99(1):3-8. doi: 10.1093/aob/mcl236. Epub 2006 Nov 10. Ann Bot. 2007. PMID: 17098753 Free PMC article. Review.
-
Regulation of malate metabolism in grape berry and other developing fruits.Phytochemistry. 2009 Jul-Aug;70(11-12):1329-44. doi: 10.1016/j.phytochem.2009.08.006. Epub 2009 Sep 15. Phytochemistry. 2009. PMID: 19762054 Review.
Cited by
-
Fruit Salad in the Lab: Comparing Botanical Species to Help Deciphering Fruit Primary Metabolism.Front Plant Sci. 2019 Jul 9;10:836. doi: 10.3389/fpls.2019.00836. eCollection 2019. Front Plant Sci. 2019. PMID: 31354750 Free PMC article. Review.
-
Differential transcriptional regulation of L-ascorbic acid content in peel and pulp of citrus fruits during development and maturation.Planta. 2014 May;239(5):1113-28. doi: 10.1007/s00425-014-2044-z. Epub 2014 Feb 25. Planta. 2014. PMID: 24567029
-
Transcript and metabolite analysis in Trincadeira cultivar reveals novel information regarding the dynamics of grape ripening.BMC Plant Biol. 2011 Nov 2;11:149. doi: 10.1186/1471-2229-11-149. BMC Plant Biol. 2011. PMID: 22047180 Free PMC article.
-
Allelic variation in paralogs of GDP-L-galactose phosphorylase is a major determinant of vitamin C concentrations in apple fruit.Plant Physiol. 2012 Nov;160(3):1613-29. doi: 10.1104/pp.112.203786. Epub 2012 Sep 21. Plant Physiol. 2012. PMID: 23001142 Free PMC article.
-
Transcriptome comparison analyses in UV-B induced AsA accumulation of Lactuca sativa L.BMC Genomics. 2023 Feb 3;24(1):61. doi: 10.1186/s12864-023-09133-7. BMC Genomics. 2023. PMID: 36737693 Free PMC article.
References
-
- Smirnoff N. Ascorbic acid: metabolism and functions of a multi-facetted molecule. Curr Opin Plant Biol. 2000;3(3):229–235. - PubMed
-
- De Gara L. In: Vitamin C Functions and Biochemistry in Animals and Plants. Asard H, May JM, Smirnoff N, editor. Vol. 1. Oxford: BIOS Scientific Publishers; 2004. Ascorbate and plant growth: from germination to cell death; pp. 83–95.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
