

An Arabidopsis flavonoid transporter is required for anther dehiscence and pollen development
- PMID: 19995827
- PMCID: PMC2803208
- DOI: 10.1093/jxb/erp312
An Arabidopsis flavonoid transporter is required for anther dehiscence and pollen development
Abstract
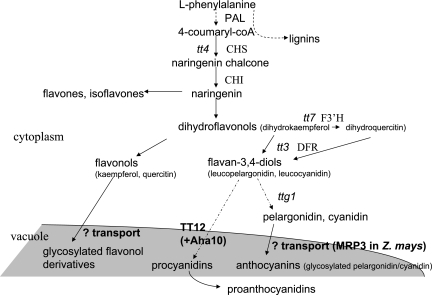
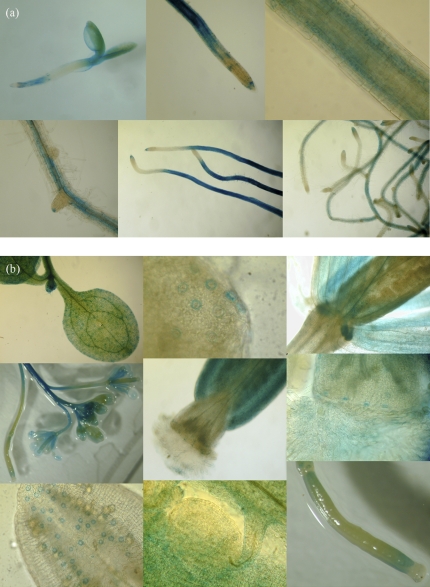
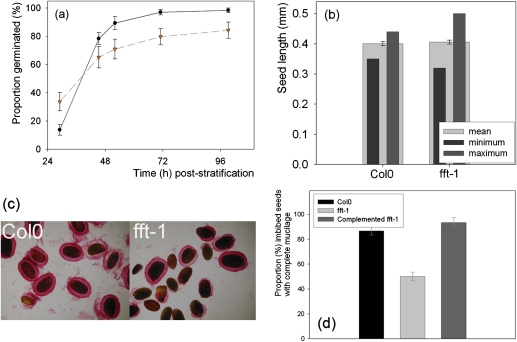
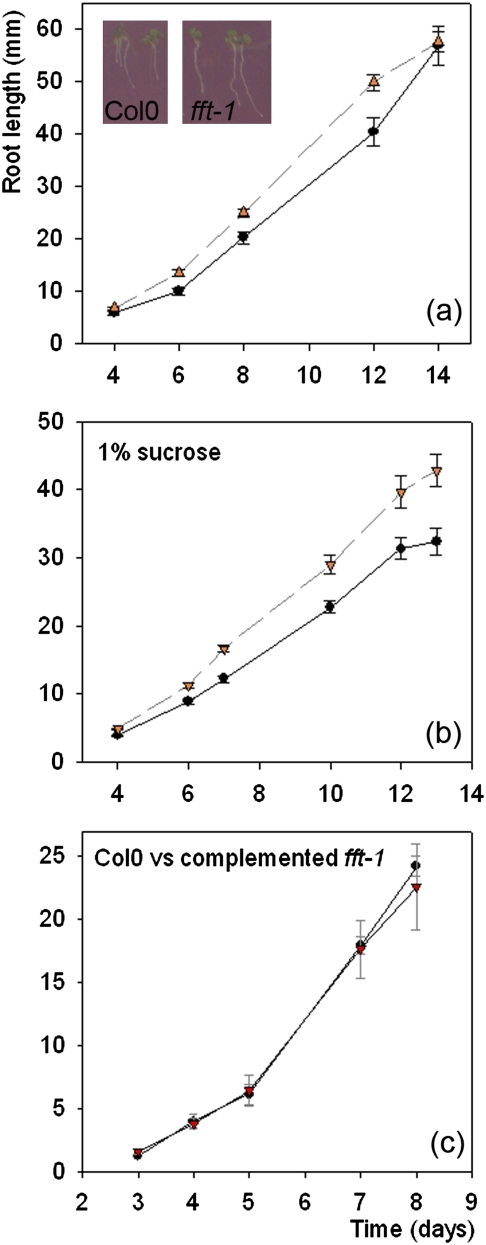
FLOWER FLAVONOID TRANSPORTER (FFT) encodes a multidrug and toxin efflux family transporter in Arabidopsis thaliana. FFT (AtDTX35) is highly transcribed in floral tissues, the transcript being localized to epidermal guard cells, including those of the anthers, stigma, siliques and nectaries. Mutant analysis demonstrates that the absence of FFT transcript affects flavonoid levels in the plant and that the altered flavonoid metabolism has wide-ranging consequences. Root growth, seed development and germination, and pollen development, release and viability are all affected. Spectrometry of mutant versus wild-type flowers shows altered levels of a glycosylated flavonol whereas anthocyanin seems unlikely to be the substrate as previously speculated. Thus, as well as adding FFT to the incompletely described flavonoid transport network, it is found that correct reproductive development in Arabidopsis is perturbed when this particular transporter is missing.
Figures
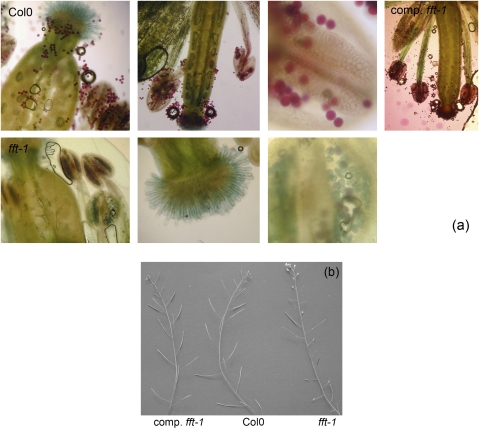
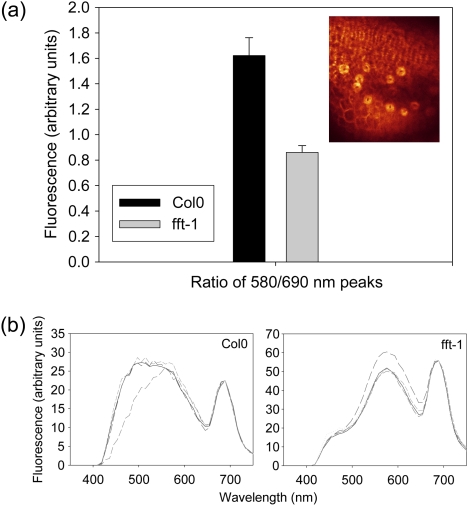
Comment in
-
Identifying the transporters of different flavonoids in plants.Plant Signal Behav. 2010 Jul;5(7):860-3. doi: 10.4161/psb.5.7.11894. Epub 2010 Jul 1. Plant Signal Behav. 2010. PMID: 20505354 Free PMC article.
Similar articles
-
Identification of an Arabidopsis plasma membrane-located ATP transporter important for anther development.Plant Cell. 2011 May;23(5):1932-44. doi: 10.1105/tpc.111.084574. Epub 2011 May 3. Plant Cell. 2011. PMID: 21540435 Free PMC article.
-
Nodulin Intrinsic Protein 7;1 Is a Tapetal Boric Acid Channel Involved in Pollen Cell Wall Formation.Plant Physiol. 2018 Nov;178(3):1269-1283. doi: 10.1104/pp.18.00604. Epub 2018 Sep 28. Plant Physiol. 2018. PMID: 30266747 Free PMC article.
-
An Arabidopsis F-box protein regulates tapetum degeneration and pollen maturation during anther development.Planta. 2010 Jul;232(2):353-66. doi: 10.1007/s00425-010-1178-x. Epub 2010 May 11. Planta. 2010. PMID: 20458496
-
The final split: the regulation of anther dehiscence.J Exp Bot. 2011 Mar;62(5):1633-49. doi: 10.1093/jxb/err014. Epub 2011 Feb 16. J Exp Bot. 2011. PMID: 21325605 Review.
-
Transcriptional regulation of anther development in Arabidopsis.Gene. 2019 Mar 20;689:202-209. doi: 10.1016/j.gene.2018.12.022. Epub 2018 Dec 17. Gene. 2019. PMID: 30572098 Review.
Cited by
-
How exposure to ALS-inhibiting gametocide tribenuron-methyl induces male sterility in rapeseed.BMC Plant Biol. 2019 Apr 2;19(1):124. doi: 10.1186/s12870-019-1722-1. BMC Plant Biol. 2019. PMID: 30940071 Free PMC article.
-
ALA-Induced Flavonols Accumulation in Guard Cells Is Involved in Scavenging H2O2 and Inhibiting Stomatal Closure in Arabidopsis Cotyledons.Front Plant Sci. 2016 Nov 15;7:1713. doi: 10.3389/fpls.2016.01713. eCollection 2016. Front Plant Sci. 2016. PMID: 27895660 Free PMC article.
-
Rice β-Glucosidase 4 (Os1βGlu4) Regulates the Hull Pigmentation via Accumulation of Salicylic Acid.Int J Mol Sci. 2022 Sep 13;23(18):10646. doi: 10.3390/ijms231810646. Int J Mol Sci. 2022. PMID: 36142555 Free PMC article.
-
Multifaceted roles of flavonoids mediating plant-microbe interactions.Microbiome. 2022 Dec 16;10(1):233. doi: 10.1186/s40168-022-01420-x. Microbiome. 2022. PMID: 36527160 Free PMC article. Review.
-
Jasmonate signaling involves the abscisic acid receptor PYL4 to regulate metabolic reprogramming in Arabidopsis and tobacco.Proc Natl Acad Sci U S A. 2011 Apr 5;108(14):5891-6. doi: 10.1073/pnas.1103010108. Epub 2011 Mar 21. Proc Natl Acad Sci U S A. 2011. PMID: 21436041 Free PMC article.
References
-
- Aarts MG, Hodge R, Kalantidis K, Florack D, Wilson ZA, Mulligan BJ, Stiekema WJ, Scott R, Pereira A. The Arabidopsis MALE STERILITY 2 protein shares similarity with reductases in elongation/condensation complexes. The Plant Journal. 1997;12:615–623. - PubMed
-
- Blankenship RE. Molecular mechanisms of photosynthesis. Oxford: Blackwell Science; 2002.
-
- Bolhar-Nordenkampf HR, Long SP, Baker NR, Oquist U, Schreiber U, Lechner EG. Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: a review of current instrumentation. Functional Ecology. 1989;3:497–514.
-
- Broun P. Transcriptional control of flavonoid biosynthesis: a complex network of conserved regulators involved in multiple aspects of differentiation in Arabidopsis. Current Opinion in Plant Biology. 2005;8:272–279. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
