

A chemical genetic approach reveals that p38alpha MAPK activation by diphosphorylation aggravates myocardial infarction and is prevented by the direct binding of SB203580
- PMID: 19996096
- PMCID: PMC2823430
- DOI: 10.1074/jbc.M109.079228
A chemical genetic approach reveals that p38alpha MAPK activation by diphosphorylation aggravates myocardial infarction and is prevented by the direct binding of SB203580
Abstract
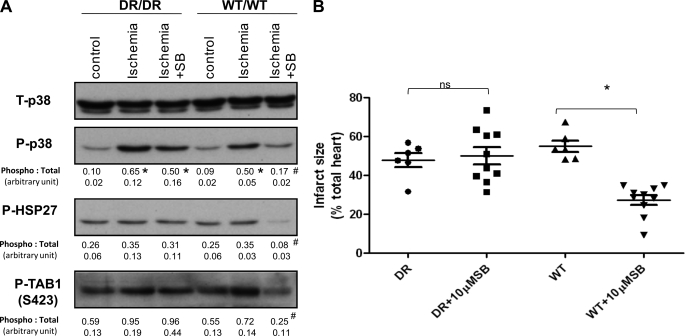
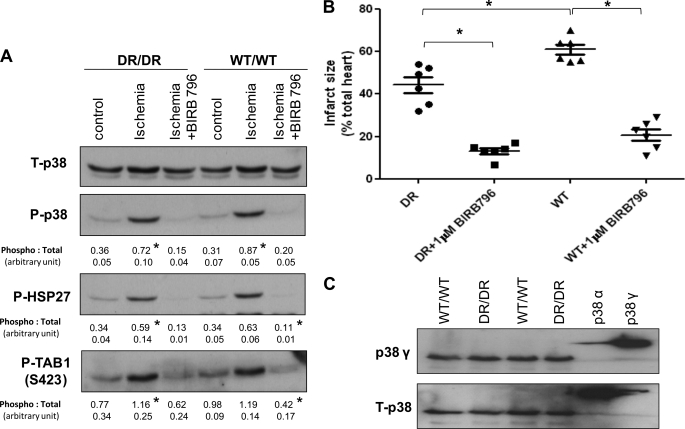
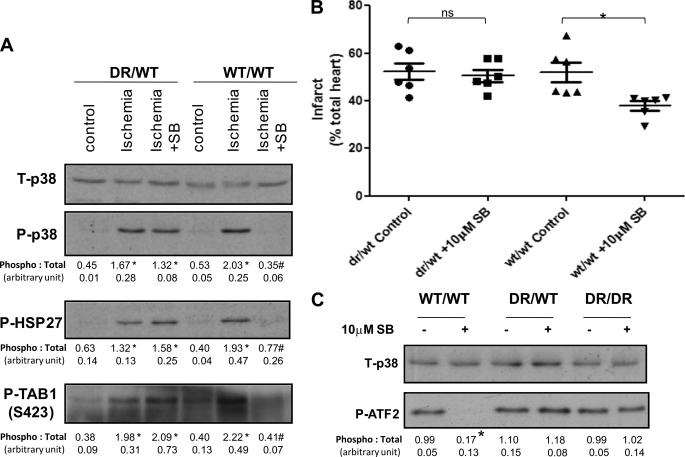
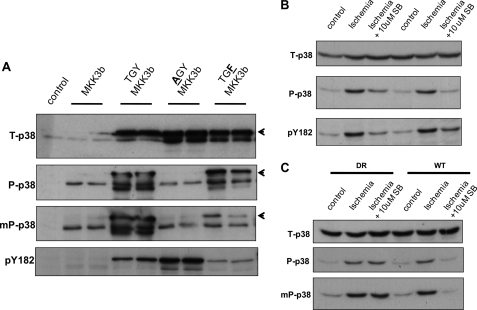
The use of nonselective pharmacological inhibitors has resulted in controversy regarding the mechanism and consequences of p38 activation during myocardial infarction. Classic p38 inhibitors such as SB203580 rely on a critical "gatekeeper" threonine residue for binding. We addressed these controversies by using mice in which the p38alpha alleles were targeted to cause substitution of the gatekeeper residue and resistance to inhibition. In homozygous drug-resistant compared with wild-type hearts, SB203580 failed to inhibit the activating phosphorylation of p38 or to reduce the infarction caused by myocardial ischemia. However, BIRB796, a p38 inhibitor not reliant on the gatekeeper for binding, similarly reduced p38-activating phosphorylation and infarction in both wild-type and knock-in mice, thereby excluding a nonspecific inhibitor-dependent phenotype resulting from the targeting strategy. Furthermore, the activation during myocardial ischemia involved phosphorylation of both the threonine and tyrosine residues in the activation loop of p38 despite the phosphorylation of the threonine alone being sufficient to create the epitope for dual phosphospecific antibody binding. Finally, SB203580 failed to reduce infarction in heterozygous drug-resistant hearts, suggesting that near complete inhibition of p38alpha kinase activity is necessary to elicit protection. These results indicate that, during myocardial ischemia, p38alpha (i) is the dominant-active p38 isoform, (ii) contributes to infarction, (iii) is responsible for the cardioprotective effect of SB203580, and (iv) is activated by a mechanism consistent with autodiphosphorylation despite this necessitating the phosphorylation of a tyrosine residue by an archetypal serine/threonine kinase.
Figures
Similar articles
-
Diverse mechanisms of myocardial p38 mitogen-activated protein kinase activation: evidence for MKK-independent activation by a TAB1-associated mechanism contributing to injury during myocardial ischemia.Circ Res. 2003 Aug 8;93(3):254-61. doi: 10.1161/01.RES.0000083490.43943.85. Epub 2003 Jun 26. Circ Res. 2003. PMID: 12829618
-
The role of RIP2 in p38 MAPK activation in the stressed heart.J Biol Chem. 2008 May 2;283(18):11964-71. doi: 10.1074/jbc.M707750200. Epub 2008 Feb 29. J Biol Chem. 2008. PMID: 18310079 Free PMC article.
-
The activation of p38 alpha, and not p38 beta, mitogen-activated protein kinase is required for ischemic preconditioning.J Mol Cell Cardiol. 2010 Jun;48(6):1324-8. doi: 10.1016/j.yjmcc.2010.02.013. Epub 2010 Feb 25. J Mol Cell Cardiol. 2010. PMID: 20188737 Free PMC article.
-
MAPKAPK-2 modulates p38-MAPK localization and small heat shock protein phosphorylation but does not mediate the injury associated with p38-MAPK activation during myocardial ischemia.Cell Stress Chaperones. 2009 Sep;14(5):477-89. doi: 10.1007/s12192-009-0101-5. Epub 2009 Feb 13. Cell Stress Chaperones. 2009. PMID: 19214782 Free PMC article.
-
The role of differential activation of p38-mitogen-activated protein kinase in preconditioned ventricular myocytes.FASEB J. 2000 Nov;14(14):2237-46. doi: 10.1096/fj.99-0671com. FASEB J. 2000. PMID: 11053245
Cited by
-
Mechanism and consequence of the autoactivation of p38α mitogen-activated protein kinase promoted by TAB1.Nat Struct Mol Biol. 2013 Oct;20(10):1182-90. doi: 10.1038/nsmb.2668. Epub 2013 Sep 15. Nat Struct Mol Biol. 2013. PMID: 24037507 Free PMC article.
-
The TAB1-p38α complex aggravates myocardial injury and can be targeted by small molecules.JCI Insight. 2018 Aug 23;3(16):e121144. doi: 10.1172/jci.insight.121144. eCollection 2018 Aug 23. JCI Insight. 2018. PMID: 30135318 Free PMC article.
-
The p38 mitogen-activated protein kinase pathway--a potential target for intervention in infarction, hypertrophy, and heart failure.J Mol Cell Cardiol. 2011 Oct;51(4):485-90. doi: 10.1016/j.yjmcc.2010.10.021. Epub 2010 Nov 6. J Mol Cell Cardiol. 2011. PMID: 21062627 Free PMC article. Review.
-
Role of p38 inhibition in cardiac ischemia/reperfusion injury.Eur J Clin Pharmacol. 2012 May;68(5):513-24. doi: 10.1007/s00228-011-1193-2. Epub 2011 Dec 29. Eur J Clin Pharmacol. 2012. PMID: 22205273 Review.
-
Endothelial-Cell-Derived Human Secretory Leukocyte Protease Inhibitor (SLPI) Protects Cardiomyocytes against Ischemia/Reperfusion Injury.Biomolecules. 2019 Oct 31;9(11):678. doi: 10.3390/biom9110678. Biomolecules. 2019. PMID: 31683729 Free PMC article.
References
-
- Ge B., Gram H., Di Padova F., Huang B., New L., Ulevitch R. J., Luo Y., Han J. (2002) Science 295, 1291–1294 - PubMed
-
- Tanno M., Bassi R., Gorog D. A., Saurin A. T., Jiang J., Heads R. J., Martin J. L., Davis R. J., Flavell R. A., Marber M. S. (2003) Circ. Res. 93, 254–261 - PubMed
-
- Fiedler B., Feil R., Hofmann F., Willenbockel C., Drexler H., Smolenski A., Lohmann S. M., Wollert K. C. (2006) J. Biol. Chem. 281, 32831–32840 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
