

Biophysical model for gamma rhythms in the olfactory bulb via subthreshold oscillations
- PMID: 19996171
- PMCID: PMC2799880
- DOI: 10.1073/pnas.0910964106
Biophysical model for gamma rhythms in the olfactory bulb via subthreshold oscillations
Abstract
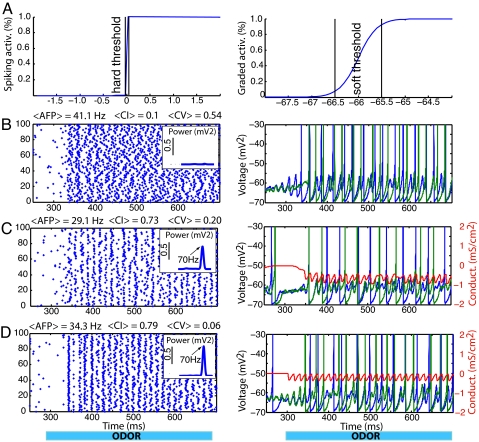
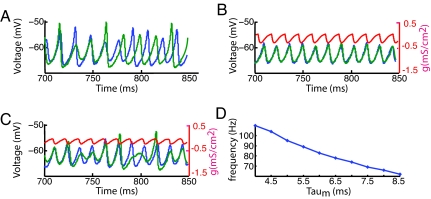
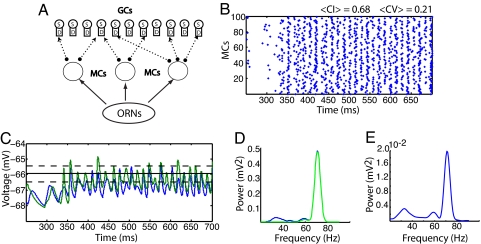
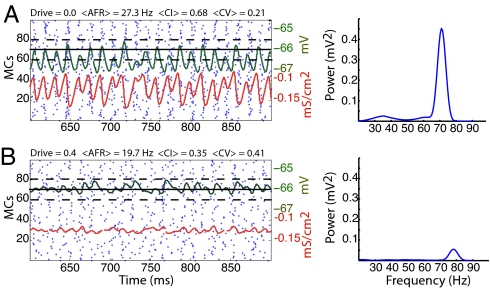
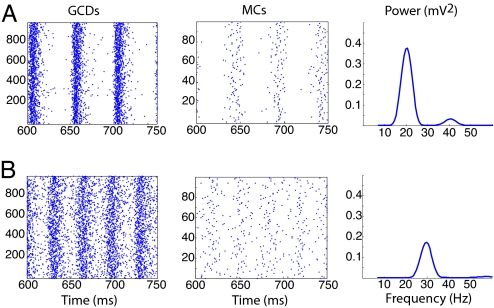
Gamma oscillations in the olfactory bulb can be produced as an interaction of subthreshold oscillations (STOs) in the mitral cells (MCs) with inhibitory granule cells (GCs). The mechanism does not require that the GCs spike, and we work in a regime in which the MCs fire at rates lower than the fast gamma rhythm they create. The frequency of the network is that of the STOs, allowing the gamma to be modulated in amplitude with only small changes in frequency. Gamma oscillations could also be obtained with spiking GCs, but only for GCs firing close to population rate. Our mechanism differs from the more standard description of the gamma oscillation, in which the the decay time of the inhibitory cells is critical to the frequency of the network.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
In vivo beta and gamma subthreshold oscillations in rat mitral cells: origin and gating by respiratory dynamics.J Neurophysiol. 2018 Jan 1;119(1):274-289. doi: 10.1152/jn.00053.2017. Epub 2017 Oct 11. J Neurophysiol. 2018. PMID: 29021388
-
Granule cell excitability regulates gamma and beta oscillations in a model of the olfactory bulb dendrodendritic microcircuit.J Neurophysiol. 2016 Aug 1;116(2):522-39. doi: 10.1152/jn.00988.2015. Epub 2016 Apr 27. J Neurophysiol. 2016. PMID: 27121582 Free PMC article.
-
Sniff rhythm-paced fast and slow gamma-oscillations in the olfactory bulb: relation to tufted and mitral cells and behavioral states.J Neurophysiol. 2013 Oct;110(7):1593-9. doi: 10.1152/jn.00379.2013. Epub 2013 Jul 17. J Neurophysiol. 2013. PMID: 23864376
-
Intraglomerular gap junctions enhance interglomerular synchrony in a sparsely connected olfactory bulb network.J Physiol. 2017 Sep 1;595(17):5965-5986. doi: 10.1113/JP274408. Epub 2017 Jul 23. J Physiol. 2017. PMID: 28640508 Free PMC article.
-
A two-layer biophysical model of cholinergic neuromodulation in olfactory bulb.J Neurosci. 2013 Feb 13;33(7):3037-58. doi: 10.1523/JNEUROSCI.2831-12.2013. J Neurosci. 2013. PMID: 23407960 Free PMC article.
Cited by
-
Top-Down Control of Inhibitory Granule Cells in the Main Olfactory Bulb Reshapes Neural Dynamics Giving Rise to a Diversity of Computations.Front Comput Neurosci. 2020 Jul 13;14:59. doi: 10.3389/fncom.2020.00059. eCollection 2020. Front Comput Neurosci. 2020. PMID: 32765248 Free PMC article.
-
A model of cholinergic modulation in olfactory bulb and piriform cortex.J Neurophysiol. 2013 Mar;109(5):1360-77. doi: 10.1152/jn.00577.2012. Epub 2012 Dec 5. J Neurophysiol. 2013. PMID: 23221406 Free PMC article.
-
Interlayer Connectivity Affects the Coherence Resonance and Population Activity Patterns in Two-Layered Networks of Excitatory and Inhibitory Neurons.Front Comput Neurosci. 2022 Apr 18;16:885720. doi: 10.3389/fncom.2022.885720. eCollection 2022. Front Comput Neurosci. 2022. PMID: 35521427 Free PMC article.
-
Cell and circuit origins of fast network oscillations in the mammalian main olfactory bulb.Elife. 2021 Oct 18;10:e74213. doi: 10.7554/eLife.74213. Elife. 2021. PMID: 34658333 Free PMC article.
-
Neuropsychiatric consequences of COVID-19 related olfactory dysfunction: could non-olfactory cortical-bound inputs from damaged olfactory bulb also contribute to cognitive impairment?Front Neurosci. 2023 Jun 22;17:1164042. doi: 10.3389/fnins.2023.1164042. eCollection 2023. Front Neurosci. 2023. PMID: 37425004 Free PMC article. No abstract available.
References
-
- Balu R, Larimer P, Strowbridge BW. Phasic stimuli evoke precisely timed spikes in intermittently discharging mitral cells. J Neurophysiol. 2004;92:743–753. - PubMed
-
- Whittington MA, Traub RD, Kopell N, Ermentrout B, Buhl EH. Inhibition-based rhythms: Experimental and mathematical observations on network dynamics. Int J Psychophysiol. 2000;38:315–336. - PubMed
-
- Börgers C, Kopell N. Synchronization in networks of excitatory and inhibitory neurons with sparse, random connectivity. Neural Comput. 2003;15:509–538. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
