

Letting Escherichia coli teach me about genome engineering
- PMID: 19996374
- PMCID: PMC2787414
- DOI: 10.1534/genetics.109.110007
Letting Escherichia coli teach me about genome engineering
Abstract
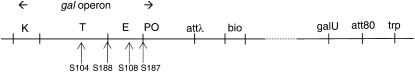
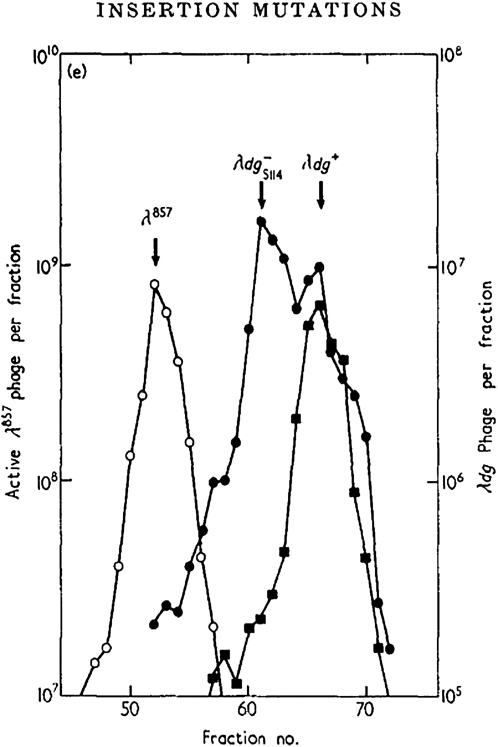
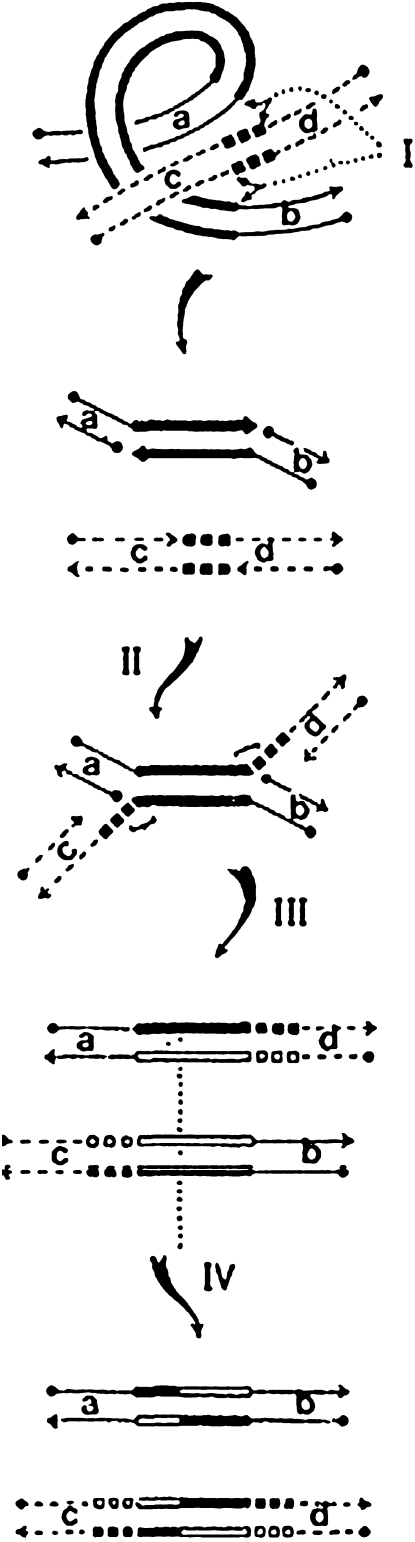
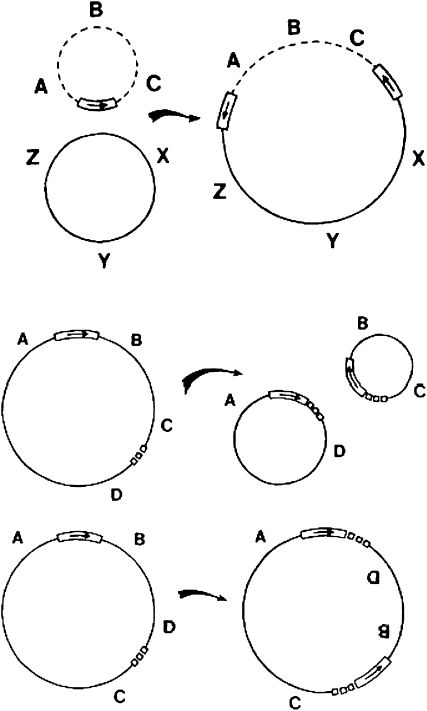
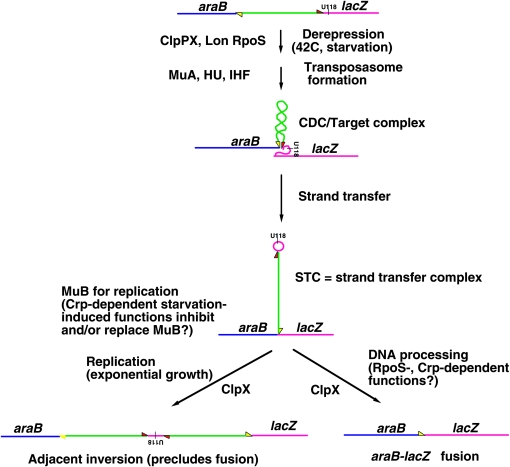
A career of following unplanned observations has serendipitously led to a deep appreciation of the capacity that bacterial cells have for restructuring their genomes in a biologically responsive manner. Routine characterization of spontaneous mutations in the gal operon guided the discovery that bacteria transpose DNA segments into new genome sites. A failed project to fuse lambda sequences to a lacZ reporter ultimately made it possible to demonstrate how readily Escherichia coli generated rearrangements necessary for in vivo cloning of chromosomal fragments into phage genomes. Thinking about the molecular mechanism of IS1 and phage Mu transposition unexpectedly clarified how transposable elements mediate large-scale rearrangements of the bacterial genome. Following up on lab lore about long delays needed to obtain Mu-mediated lacZ protein fusions revealed a striking connection between physiological stress and activation of DNA rearrangement functions. Examining the fate of Mudlac DNA in sectored colonies showed that these same functions are subject to developmental control, like controlling elements in maize. All these experiences confirmed Barbara McClintock's view that cells frequently respond to stimuli by restructuring their genomes and provided novel insights into the natural genetic engineering processes involved in evolution.
Figures
References
-
- Beckwith, J. R., E. R. Signer and W. Epstein, 1966. Transposition of the Lac region of E. coli. Cold Spring Harb. Symp. Quant. Biol. 31 393–401. - PubMed
-
- Bennetzen, J. L., 2005. Transposable elements, gene creation and genome rearrangement in flowering plants. Curr. Opin. Genet. Dev. 15 621–627. - PubMed
-
- Bjedov, I., O. Tenaillon, B. Gérard, V. Souza, E. Denamur et al., 2003. Stress-induced mutagenesis in bacteria. Science 300 1404–1409. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
