

Homeobox transcription factors are required for conidiation and appressorium development in the rice blast fungus Magnaporthe oryzae
- PMID: 19997500
- PMCID: PMC2779367
- DOI: 10.1371/journal.pgen.1000757
Homeobox transcription factors are required for conidiation and appressorium development in the rice blast fungus Magnaporthe oryzae
Abstract
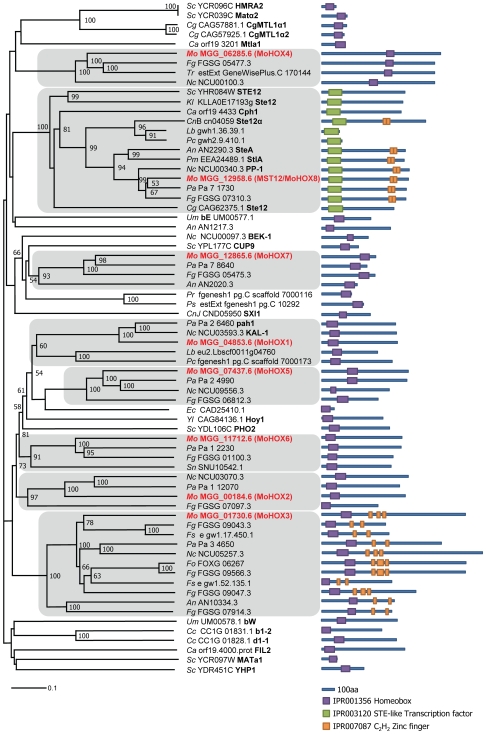
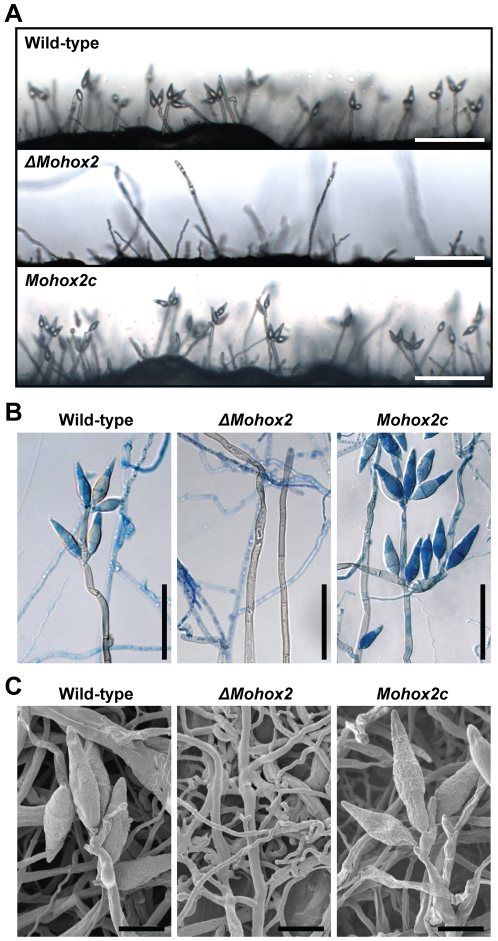
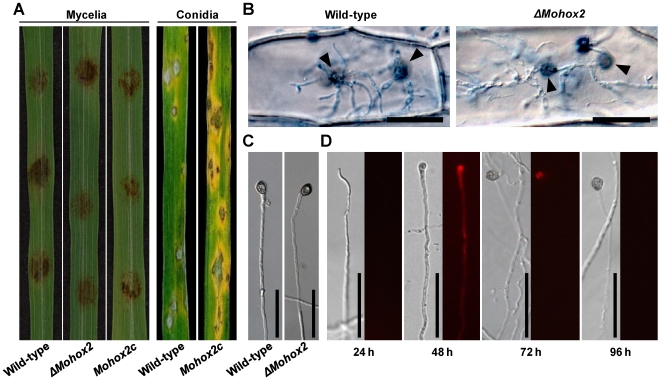
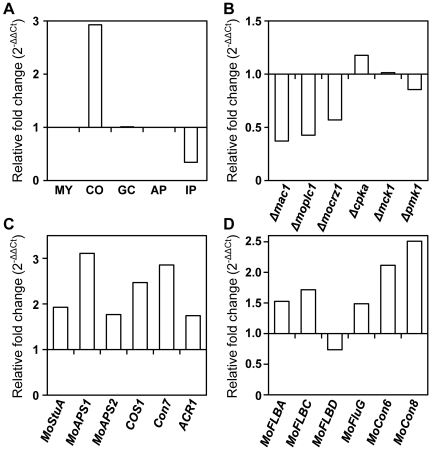
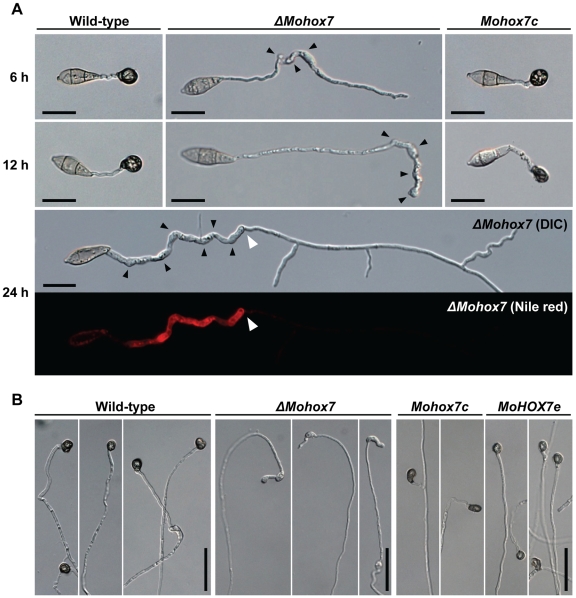
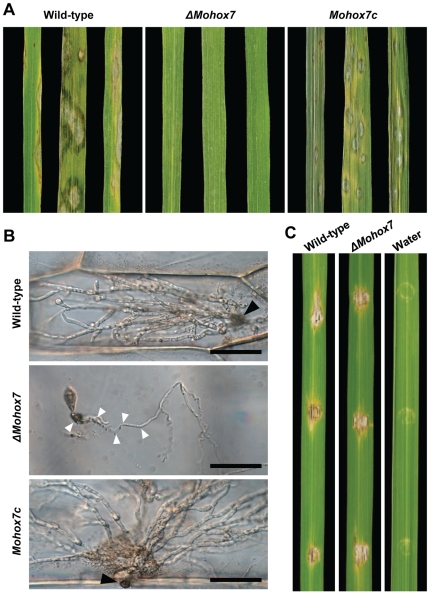
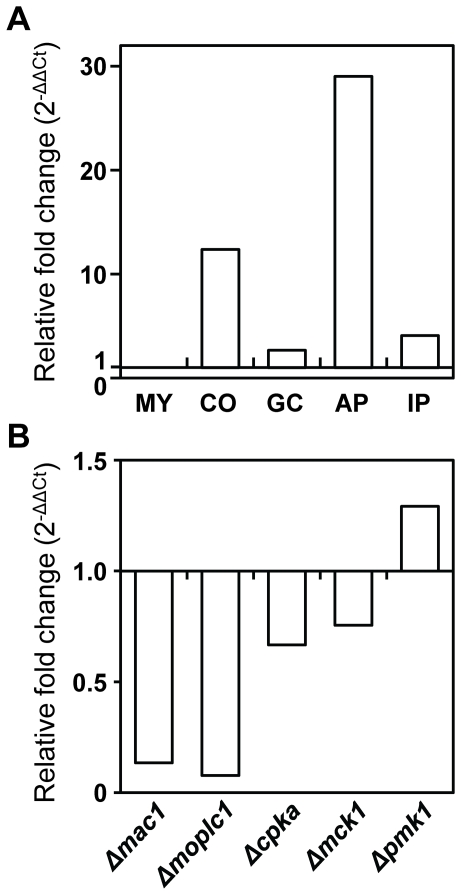
The appropriate development of conidia and appressoria is critical in the disease cycle of many fungal pathogens, including Magnaporthe oryzae. A total of eight genes (MoHOX1 to MoHOX8) encoding putative homeobox transcription factors (TFs) were identified from the M. oryzae genome. Knockout mutants for each MoHOX gene were obtained via homology-dependent gene replacement. Two mutants, DeltaMohox3 and DeltaMohox5, exhibited no difference to wild-type in growth, conidiation, conidium size, conidial germination, appressorium formation, and pathogenicity. However, the DeltaMohox1 showed a dramatic reduction in hyphal growth and increase in melanin pigmentation, compared to those in wild-type. DeltaMohox4 and DeltaMohox6 showed significantly reduced conidium size and hyphal growth, respectively. DeltaMohox8 formed normal appressoria, but failed in pathogenicity, probably due to defects in the development of penetration peg and invasive growth. It is most notable that asexual reproduction was completely abolished in DeltaMohox2, in which no conidia formed. DeltaMohox2 was still pathogenic through hypha-driven appressoria in a manner similar to that of the wild-type. However, DeltaMohox7 was unable to form appressoria either on conidial germ tubes, or at hyphal tips, being non-pathogenic. These factors indicate that M. oryzae is able to cause foliar disease via hyphal appressorium-mediated penetration, and MoHOX7 is mutually required to drive appressorium formation from hyphae and germ tubes. Transcriptional analyses suggest that the functioning of M. oryzae homeobox TFs is mediated through the regulation of gene expression and is affected by cAMP and Ca(2+) signaling and/or MAPK pathways. The divergent roles of this gene set may help reveal how the genome and regulatory pathways evolved within the rice blast pathogen and close relatives.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Ou SH. Rice Diseases. Kew, England: Commonwealth Mycological Institute; 1985. pp. 97–184.
-
- Ebbole DJ. Magnaporthe as a model for understanding host-pathogen interactions. Annu Rev Phytopathol. 2007;45:437–456. - PubMed
-
- Dean RA, Talbot NJ, Ebbole DJ, Farman ML, Mitchell TK, et al. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature. 2005;434:980–986. - PubMed
-
- Valent B. Rice blast as a model system for plant pathology. Phytopathology. 1990;80:33–36.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
