

Genome-wide identification of Schistosoma japonicum microRNAs using a deep-sequencing approach
- PMID: 19997615
- PMCID: PMC2785426
- DOI: 10.1371/journal.pone.0008206
Genome-wide identification of Schistosoma japonicum microRNAs using a deep-sequencing approach
Abstract
Background: Human schistosomiasis is one of the most prevalent and serious parasitic diseases worldwide. Schistosoma japonicum is one of important pathogens of this disease. MicroRNAs (miRNAs) are a large group of non-coding RNAs that play important roles in regulating gene expression and protein translation in animals. Genome-wide identification of miRNAs in a given organism is a critical step to facilitating our understanding of genome organization, genome biology, evolution, and posttranscriptional regulation.
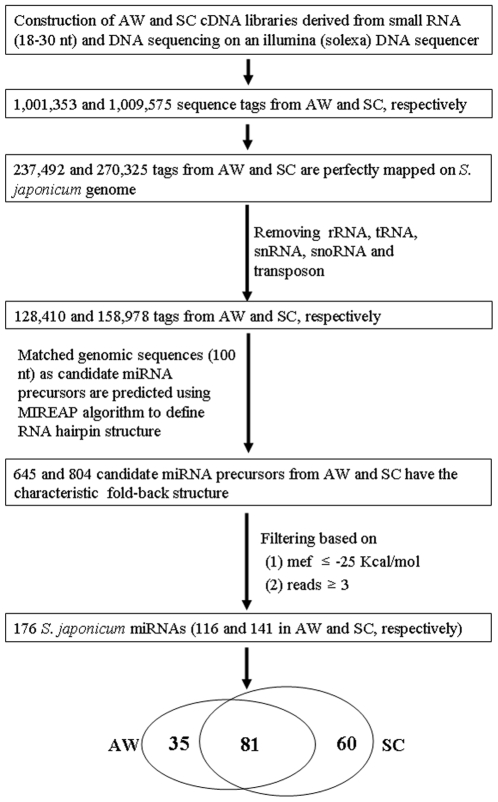
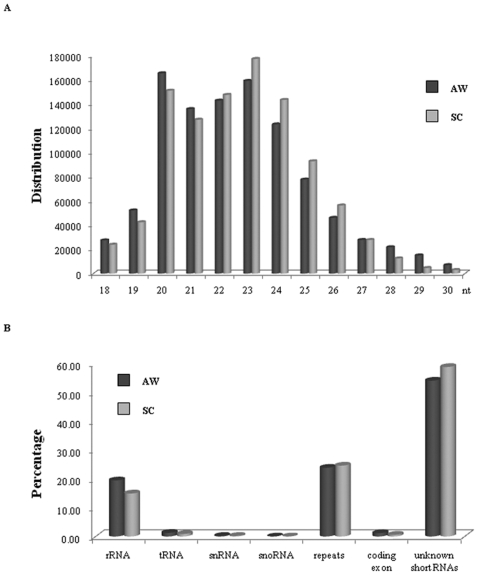
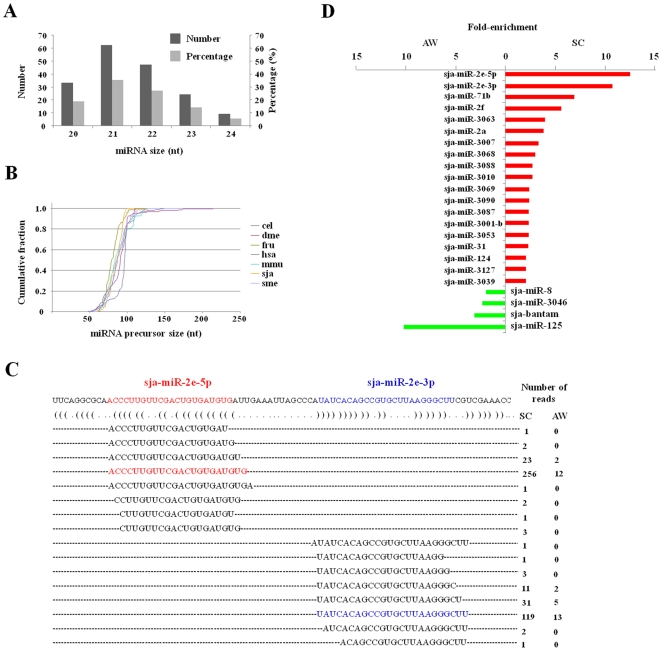
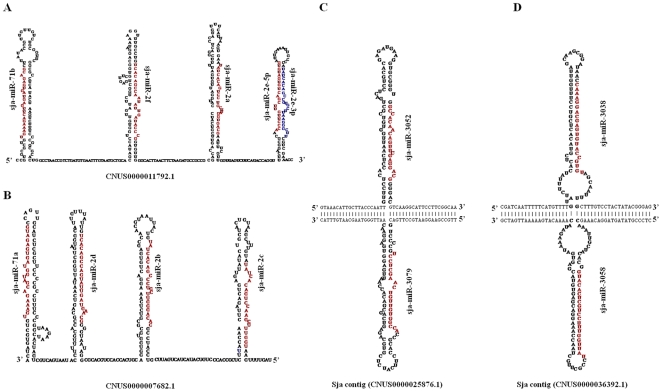
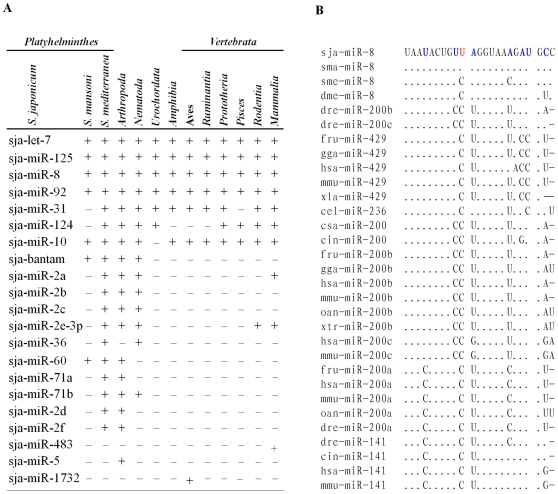
Methodology/principal findings: We sequenced two small RNA libraries prepared from different stages of the life cycle of S. japonicum, immature schistosomula and mature pairing adults, through a deep DNA sequencing approach, which yielded approximately 12 million high-quality short sequence reads containing a total of approximately 2 million non-redundant tags. Based on a bioinformatics pipeline, we identified 176 new S. japonicum miRNAs, of which some exhibited a differential pattern of expression between the two stages. Although 21 S. japonicum miRNAs are orthologs of known miRNAs within the metazoans, some nucleotides at many positions of Schistosoma miRNAs, such as miR-8, let-7, miR-10, miR-31, miR-92, miR-124, and miR-125, are indeed significantly distinct from other bilaterian orthologs. In addition, both miR-71 and some miR-2 family members in tandem are found to be clustered in a reversal direction model on two genomic loci, and two pairs of novel S. japonicum miRNAs were derived from sense and antisense DNA strands at the same genomic loci.
Conclusions/significance: The collection of S. japonicum miRNAs could be used as a new platform to study the genomic structure, gene regulation and networks, evolutionary processes, development, and host-parasite interactions. Some S. japonicum miRNAs and their clusters could represent the ancestral forms of the conserved orthologues and a model for the genesis of novel miRNAs.
Conflict of interest statement
Figures
Similar articles
-
Identification and characterization of novel microRNAs from Schistosoma japonicum.PLoS One. 2008;3(12):e4034. doi: 10.1371/journal.pone.0004034. Epub 2008 Dec 24. PLoS One. 2008. PMID: 19107204 Free PMC article.
-
Identification and characterization of microRNAs and endogenous siRNAs in Schistosoma japonicum.BMC Genomics. 2010 Jan 21;11:55. doi: 10.1186/1471-2164-11-55. BMC Genomics. 2010. PMID: 20092619 Free PMC article.
-
Comparative characterization of microRNAs in Schistosoma japonicum schistosomula from Wistar rats and BALB/c mice.Parasitol Res. 2015 Jul;114(7):2639-47. doi: 10.1007/s00436-015-4468-1. Epub 2015 Apr 19. Parasitol Res. 2015. PMID: 25895062
-
High-throughput sequencing of RNAs isolated by cross-linking immunoprecipitation (HITS-CLIP) reveals Argonaute-associated microRNAs and targets in Schistosoma japonicum.Parasit Vectors. 2015 Nov 14;8:589. doi: 10.1186/s13071-015-1203-9. Parasit Vectors. 2015. PMID: 26577460 Free PMC article.
-
Comparative analysis of microRNA expression profiles of adult Schistosoma japonicum isolated from water buffalo and yellow cattle.Parasit Vectors. 2019 May 2;12(1):196. doi: 10.1186/s13071-019-3450-7. Parasit Vectors. 2019. PMID: 31046821 Free PMC article.
Cited by
-
Identification of a Schistosoma japonicum MicroRNA That Suppresses Hepatoma Cell Growth and Migration by Targeting Host FZD4 Gene.Front Cell Infect Microbiol. 2022 Jan 31;12:786543. doi: 10.3389/fcimb.2022.786543. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35174106 Free PMC article.
-
Cross-Species Suppression of Hepatoma Cell Growth and Migration by a Schistosoma japonicum MicroRNA.Mol Ther Nucleic Acids. 2019 Dec 6;18:400-412. doi: 10.1016/j.omtn.2019.09.006. Epub 2019 Sep 17. Mol Ther Nucleic Acids. 2019. PMID: 31655260 Free PMC article.
-
miR-8b is involved in brain and eye regeneration of Dugesia japonica in head regeneration.Biol Open. 2021 Jun 15;10(6):bio058538. doi: 10.1242/bio.058538. Epub 2021 Jun 29. Biol Open. 2021. PMID: 34184734 Free PMC article.
-
microRNAs in parasites and parasite infection.RNA Biol. 2013 Mar;10(3):371-9. doi: 10.4161/rna.23716. Epub 2013 Feb 7. RNA Biol. 2013. PMID: 23392243 Free PMC article. Review.
-
Dynamic Changes in the Global Transcriptome and MicroRNAome Reveal Complex miRNA-mRNA Regulation in Early Stages of the Bi-Directional Development of Echinococcus granulosus Protoscoleces.Front Microbiol. 2020 Apr 9;11:654. doi: 10.3389/fmicb.2020.00654. eCollection 2020. Front Microbiol. 2020. PMID: 32373094 Free PMC article.
References
-
- King CH, Dickman K, Tisch DJ. Reassessment of the cost of chronic helmintic infection: a meta-analysis of disability-related outcomes in endemic schistosomiasis. Lancet. 2005;365(9470):1561–1569. - PubMed
-
- Salzet M, Capron A, Stefano GB. Molecular crosstalk in host-parasite relationships: schistosome- and leech-host interactions. Parasitol Today. 2000;16(12):536–540. - PubMed
-
- Hu W, Yan Q, Shen DK, Liu F, Zhu ZD, et al. Evolutionary and biomedical implications of a Schistosoma japonicum complementary DNA resource. Nat Gene. 2003;35:139–147. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
