

Seasonal differences of gene expression profiles in song sparrow (Melospiza melodia) hypothalamus in relation to territorial aggression
- PMID: 19997634
- PMCID: PMC2780717
- DOI: 10.1371/journal.pone.0008182
Seasonal differences of gene expression profiles in song sparrow (Melospiza melodia) hypothalamus in relation to territorial aggression
Abstract
Background: Male song sparrows (Melospiza melodia) are territorial year-round; however, neuroendocrine responses to simulated territorial intrusion (STI) differ between breeding (spring) and non-breeding seasons (autumn). In spring, exposure to STI leads to increases in luteinizing hormone and testosterone, but not in autumn. These observations suggest that there are fundamental differences in the mechanisms driving neuroendocrine responses to STI between seasons. Microarrays, spotted with EST cDNA clones of zebra finch, were used to explore gene expression profiles in the hypothalamus after territorial aggression in two different seasons.
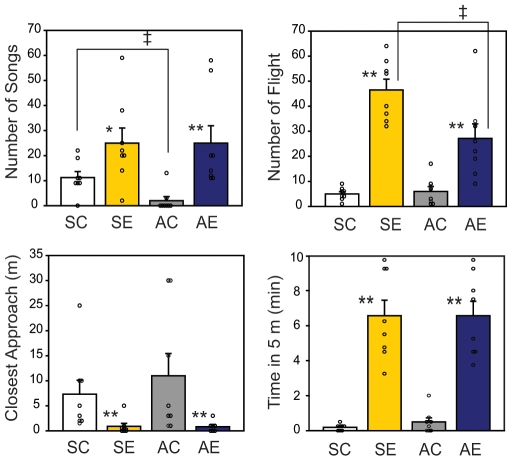
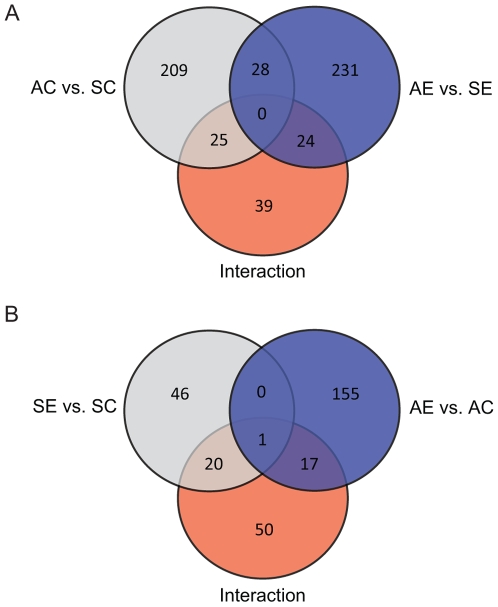
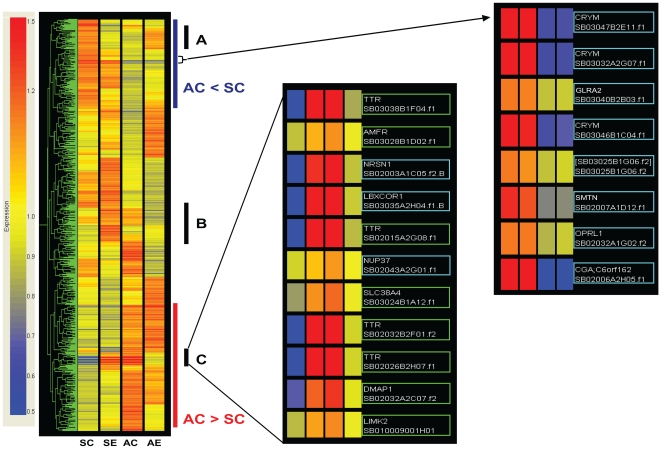
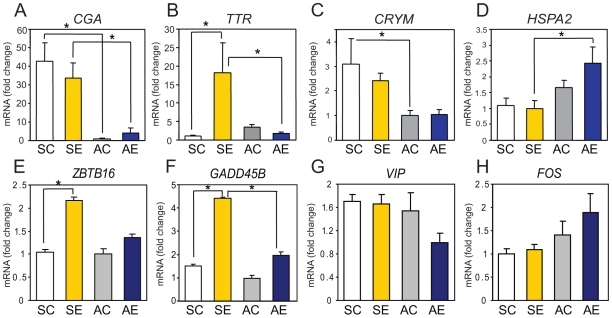
Methodology/principal findings: Free-living territorial male song sparrows were exposed to either conspecific or heterospecific (control) males in an STI in spring and autumn. Behavioral data were recorded, whole hypothalami were collected, and microarray hybridizations were performed. Quantitative PCR was performed for validation. Our results show 262 cDNAs were differentially expressed between spring and autumn in the control birds. There were 173 cDNAs significantly affected by STI in autumn; however, only 67 were significantly affected by STI in spring. There were 88 cDNAs that showed significant interactions in both season and STI.
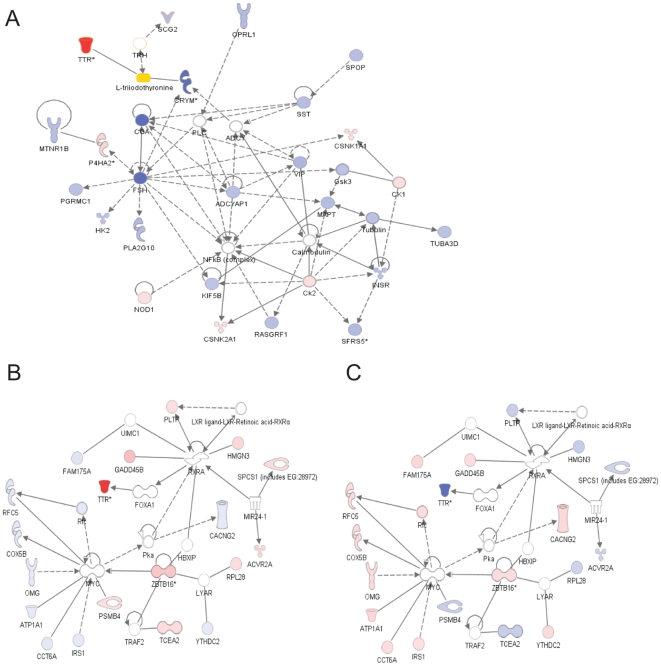
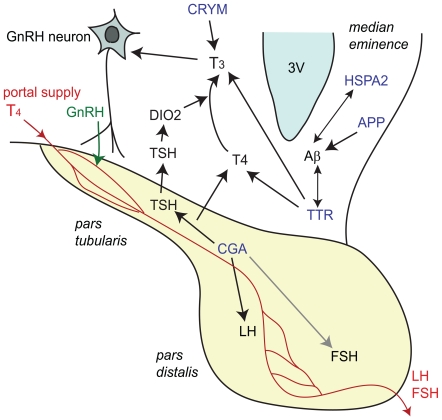
Conclusions/significance: Results suggest that STI drives differential genomic responses in the hypothalamus in the spring vs. autumn. The number of cDNAs differentially expressed in relation to season was greater than in relation to social interactions, suggesting major underlying seasonal effects in the hypothalamus which may determine the differential response upon social interaction. Functional pathway analyses implicated genes that regulate thyroid hormone action and neuroplasticity as targets of this neuroendocrine regulation.
Conflict of interest statement
Figures
Similar articles
-
Regulation of territorial behavior in the sedentary song sparrow, Melospiza melodia morphna.Horm Behav. 1994 Mar;28(1):1-15. doi: 10.1006/hbeh.1994.1001. Horm Behav. 1994. PMID: 8034278
-
The role of androgen receptors in regulating territorial aggression in male song sparrows.Horm Behav. 2010 Jan;57(1):86-95. doi: 10.1016/j.yhbeh.2009.09.015. Epub 2009 Sep 30. Horm Behav. 2010. PMID: 19799905
-
An Aggressive Interaction Rapidly Increases Brain Androgens in a Male Songbird during the Non-breeding Season.J Neurosci. 2024 Jun 5;44(23):e1095232024. doi: 10.1523/JNEUROSCI.1095-23.2024. J Neurosci. 2024. PMID: 38658166 Free PMC article.
-
Neuroendocrine Mechanisms Underlying Non-breeding Aggression: Common Strategies Between Birds and Fish.Front Neural Circuits. 2021 Jul 29;15:716605. doi: 10.3389/fncir.2021.716605. eCollection 2021. Front Neural Circuits. 2021. PMID: 34393727 Free PMC article. Review.
-
Testosterone and aggression: Berthold, birds and beyond.J Neuroendocrinol. 2006 Jul;18(7):543-51. doi: 10.1111/j.1365-2826.2006.01440.x. J Neuroendocrinol. 2006. PMID: 16774503 Free PMC article. Review.
Cited by
-
Lysine metabolism in mammalian brain: an update on the importance of recent discoveries.Amino Acids. 2013 Dec;45(6):1249-72. doi: 10.1007/s00726-013-1590-1. Epub 2013 Sep 17. Amino Acids. 2013. PMID: 24043460 Free PMC article. Review.
-
Testosterone affects neural gene expression differently in male and female juncos: a role for hormones in mediating sexual dimorphism and conflict.PLoS One. 2013 Apr 16;8(4):e61784. doi: 10.1371/journal.pone.0061784. Print 2013. PLoS One. 2013. PMID: 23613935 Free PMC article.
-
Natural variation in brain gene expression profiles of aggressive and nonaggressive individual sticklebacks.Behaviour. 2016;153(13-14):1723-1743. doi: 10.1163/1568539X-00003393. Behaviour. 2016. PMID: 29046592 Free PMC article.
-
RNA-seq transcriptome analysis of male and female zebra finch cell lines.Genomics. 2012 Dec;100(6):363-9. doi: 10.1016/j.ygeno.2012.08.002. Epub 2012 Aug 21. Genomics. 2012. PMID: 22922019 Free PMC article.
-
Transcriptional regulation of brain gene expression in response to a territorial intrusion.Proc Biol Sci. 2012 Dec 22;279(1749):4929-38. doi: 10.1098/rspb.2012.2087. Epub 2012 Oct 24. Proc Biol Sci. 2012. PMID: 23097509 Free PMC article.
References
-
- Demas GE, Moffatt CA, Drazen DL, Nelson RJ. Castration does not inhibit aggressive behavior in adult male prairie voles (Microtus ochrogaster). Physiol Behav. 1999;66:59–62. - PubMed
-
- Trainor BC, Marler CA. Testosterone, paternal behavior, and aggression in the monogamous California mouse (Peromyscus californicus). Horm Behav. 2001;40:32–42. - PubMed
-
- Wingfield JC. A continuing saga: the role of testosterone in aggression. Horm Behav. 2005;48:253–255; discussion 256–258. - PubMed
-
- Wingfield JC, Hegner RE, Dufty J, Alfred M, Ball GF. The “Challenge Hypothesis”: Theoretical Implications for Patterns of Testosterone Secretion, Mating Systems, and Breeding Strategies. The American Naturalist. 1990;136:829.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
