

Role of the signal peptide in the synthesis and processing of the glucagon-like peptide-1 receptor
- PMID: 20002095
- PMCID: PMC2823368
- DOI: 10.1111/j.1476-5381.2009.00517.x
Role of the signal peptide in the synthesis and processing of the glucagon-like peptide-1 receptor
Abstract
Background and purpose: The glucagon-like peptide-1 receptor (GLP-1R) belongs to Family B of the G protein-coupled receptor superfamily and is a target for treatment of type 2 diabetes. Family B G protein-coupled receptors contain a putative N-terminal signal peptide, but its role in receptor synthesis and trafficking are unclear. Further, the signal peptide is not cleaved in at least one family member.
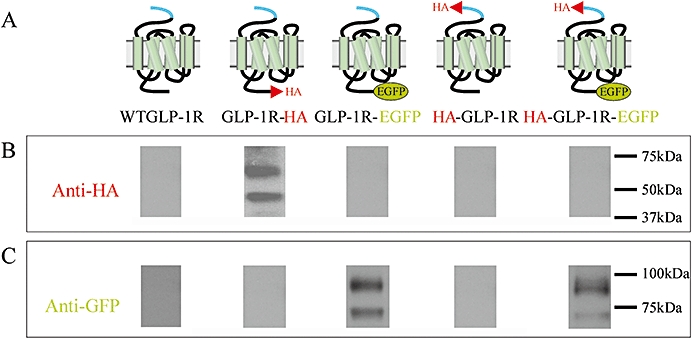
Experimental approach: We examined receptor glycosylation and the role of the signal peptide in GLP-1R synthesis and trafficking using constructs containing epitope tags at the N- and/or C-terminus and in which the signal peptide sequence was either present or absent.
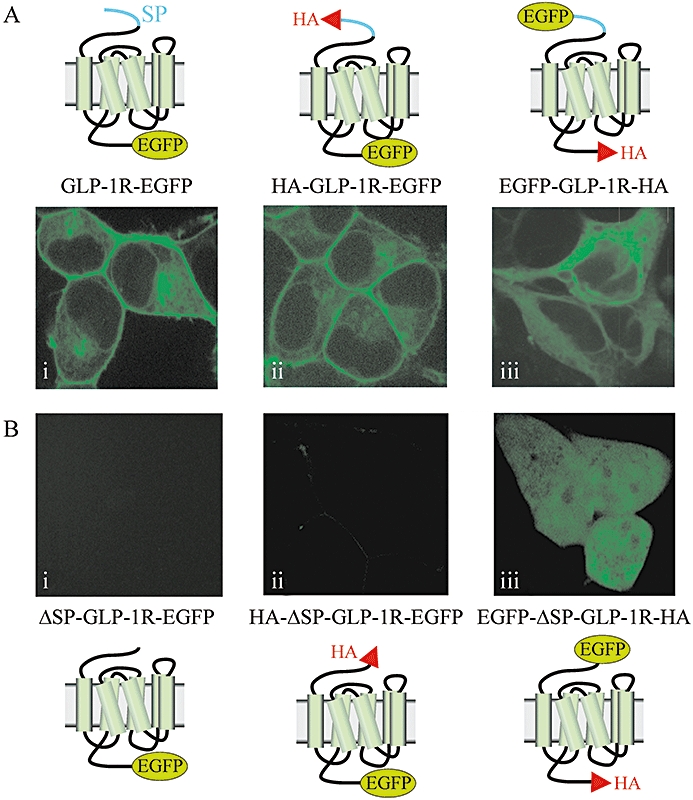
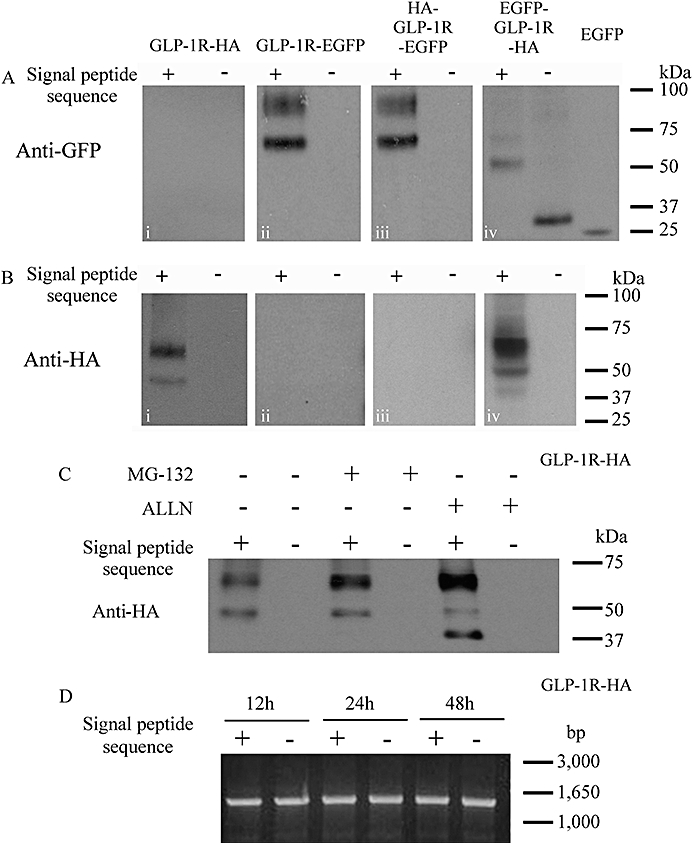
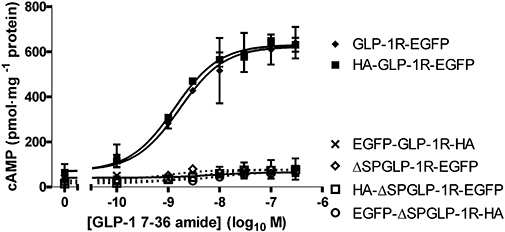
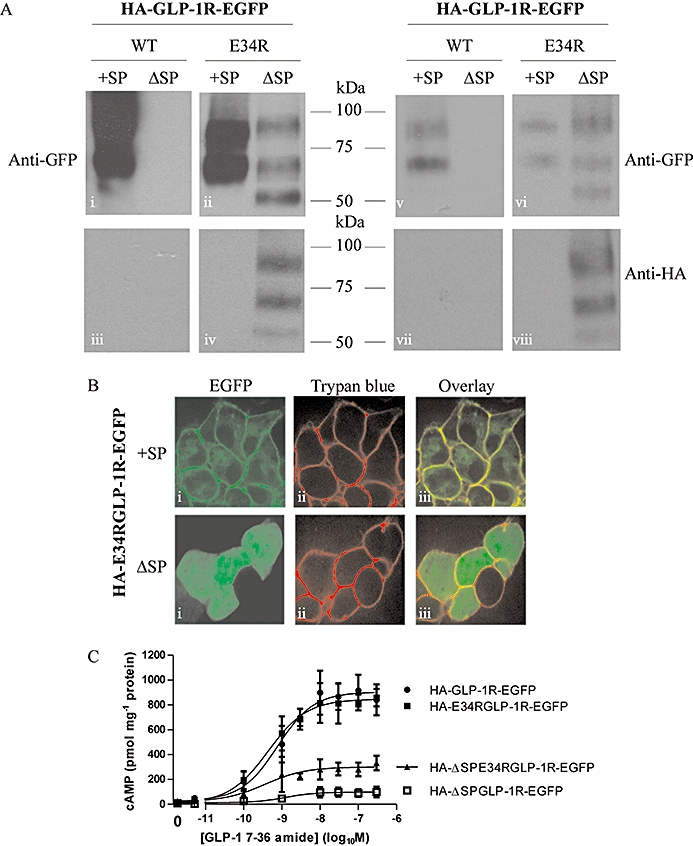
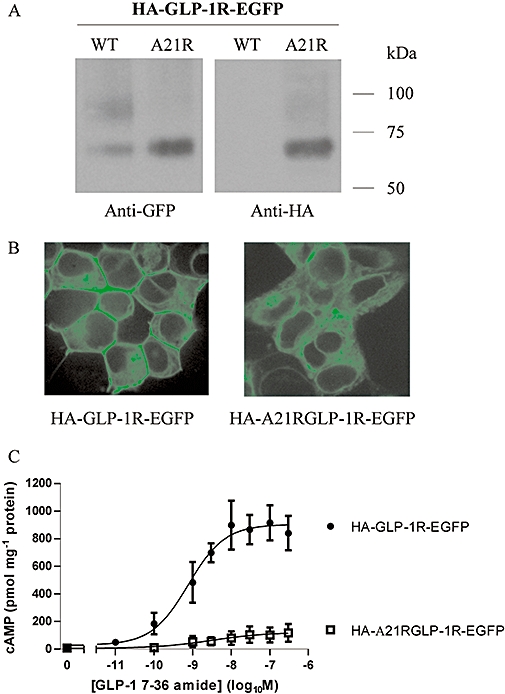
Key results: The signal peptide was absolutely required for GLP-1R synthesis but could be substituted to some extent by increasing positive charge in the N-terminal region of the receptor flanking the signal peptide. The signal peptide is cleaved during synthesis and processing of the receptor. An enhanced GFP-epitope tag at the N-terminus of the receptor permitted synthesis of the receptor but blocked signal peptide cleavage and prevented trafficking to the plasma membrane. Cleavage site mutation allowed synthesis of a full-length receptor, blocked signal peptide cleavage and caused retention within the endoplasmic reticulum.
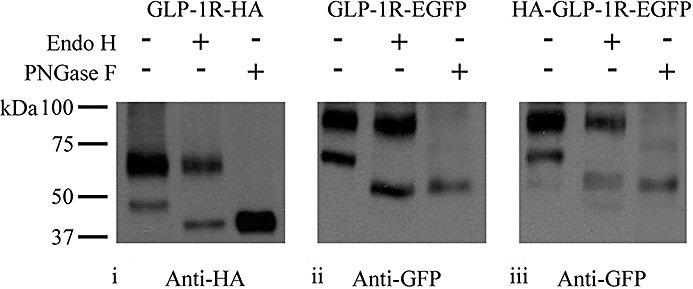
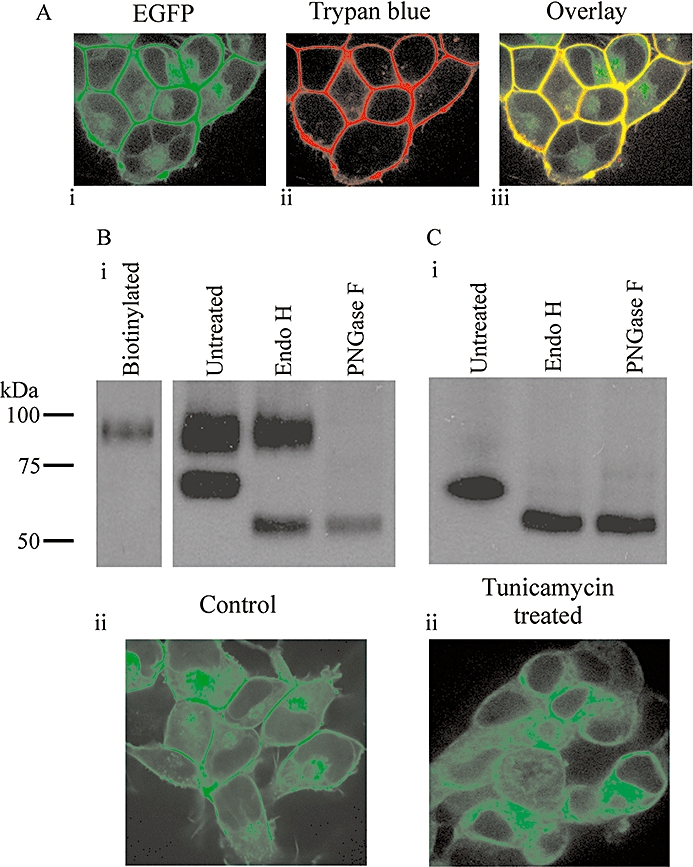
Conclusions and implications: Signal peptide cleavage was not essential for receptor synthesis but was obligatory for processing and trafficking of receptors to the plasma membrane. Further, the GLP-1R is subject to N-linked glycosylation and only the mature, fully glycosylated form of the receptor is present in the plasma membrane. Inhibition of glycosylation prevents processing and cell surface expression of the GLP-1R.
Figures

Similar articles
-
The putative signal peptide of glucagon-like peptide-1 receptor is not required for receptor synthesis but promotes receptor expression.Biosci Rep. 2014 Nov 21;34(6):e00152. doi: 10.1042/BSR20140120. Biosci Rep. 2014. PMID: 25330813 Free PMC article.
-
The regions within the N-terminus critical for human glucagon like peptide-1 receptor (hGLP-1R) cell surface expression.Sci Rep. 2014 Dec 15;4:7410. doi: 10.1038/srep07410. Sci Rep. 2014. PMID: 25502804 Free PMC article.
-
Functional consequences of glucagon-like peptide-1 receptor cross-talk and trafficking.J Biol Chem. 2015 Jan 9;290(2):1233-43. doi: 10.1074/jbc.M114.592436. Epub 2014 Dec 1. J Biol Chem. 2015. PMID: 25451942 Free PMC article.
-
Caveolin-1 regulates cellular trafficking and function of the glucagon-like Peptide 1 receptor.Mol Endocrinol. 2006 Dec;20(12):3400-11. doi: 10.1210/me.2006-0178. Epub 2006 Aug 24. Mol Endocrinol. 2006. PMID: 16931572
-
The glucagon-like peptide-2 receptor C terminus modulates beta-arrestin-2 association but is dispensable for ligand-induced desensitization, endocytosis, and G-protein-dependent effector activation.J Biol Chem. 2005 Jun 10;280(23):22124-34. doi: 10.1074/jbc.M500078200. Epub 2005 Apr 6. J Biol Chem. 2005. PMID: 15817468
Cited by
-
Prediction of Signal Peptides in Proteins from Malaria Parasites.Int J Mol Sci. 2018 Nov 22;19(12):3709. doi: 10.3390/ijms19123709. Int J Mol Sci. 2018. PMID: 30469512 Free PMC article.
-
Structural and molecular conservation of glucagon-like Peptide-1 and its receptor confers selective ligand-receptor interaction.Front Endocrinol (Lausanne). 2012 Nov 19;3:141. doi: 10.3389/fendo.2012.00141. eCollection 2012. Front Endocrinol (Lausanne). 2012. PMID: 23181056 Free PMC article.
-
Role of N-linked glycosylation in biosynthesis, trafficking, and function of the human glucagon-like peptide 1 receptor.Am J Physiol Endocrinol Metab. 2010 Jul;299(1):E62-8. doi: 10.1152/ajpendo.00067.2010. Epub 2010 Apr 20. Am J Physiol Endocrinol Metab. 2010. PMID: 20407008 Free PMC article.
-
The structure and function of the glucagon-like peptide-1 receptor and its ligands.Br J Pharmacol. 2012 May;166(1):27-41. doi: 10.1111/j.1476-5381.2011.01687.x. Br J Pharmacol. 2012. PMID: 21950636 Free PMC article. Review.
-
The putative signal peptide of glucagon-like peptide-1 receptor is not required for receptor synthesis but promotes receptor expression.Biosci Rep. 2014 Nov 21;34(6):e00152. doi: 10.1042/BSR20140120. Biosci Rep. 2014. PMID: 25330813 Free PMC article.
References
-
- Alken M, Schmidt A, Rutz C, Furkert J, Kleinau G, Rosenthal W, et al. The sequence after the signal peptide of the G protein-coupled endothelin B receptor is required for efficient translocon gating at the endoplasmic reticulum membrane. Mol Pharmacol. 2009;75:801–811. - PubMed
-
- Baggio LL, Drucker DJ. Biology of incretins: GLP-1 and GIP. Gastroenterology. 2007;132:2131–2157. - PubMed
-
- Bendtsen JD, Nielsen H, von Heijne G, Brunak S. Improved prediction of signal peptides: SignalP 3.0. J Mol Biol. 2004;340:783–795. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
