

Mechanism of primitive duct formation in the pancreas and submandibular glands: a role for SDF-1
- PMID: 20003423
- PMCID: PMC2801489
- DOI: 10.1186/1471-213X-9-66
Mechanism of primitive duct formation in the pancreas and submandibular glands: a role for SDF-1
Abstract
Background: The exocrine pancreas is composed of a branched network of ducts connected to acini. They are lined by a monolayered epithelium that derives from the endoderm and is surrounded by mesoderm-derived mesenchyme. The morphogenic mechanisms by which the ductal network is established as well as the signaling pathways involved in this process are poorly understood.
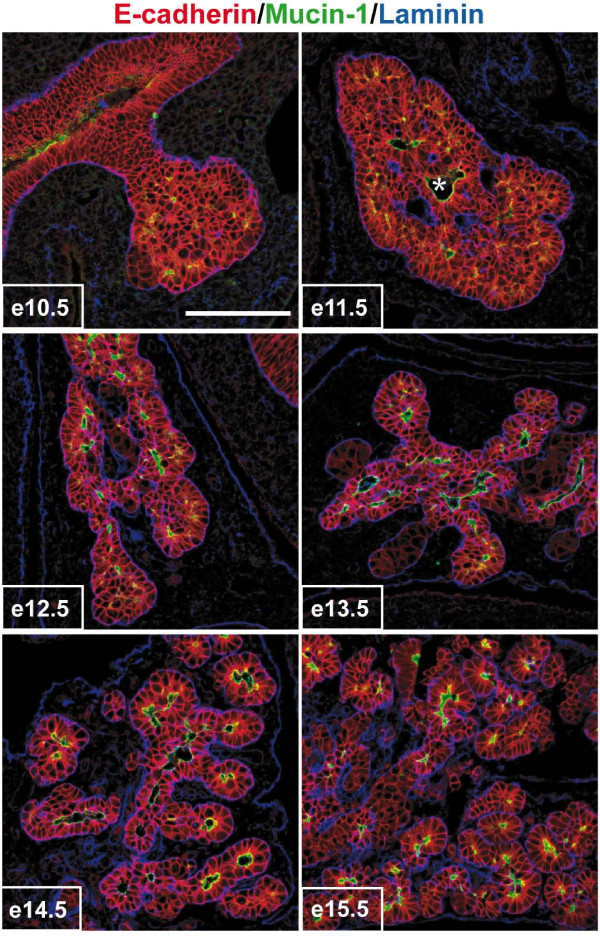
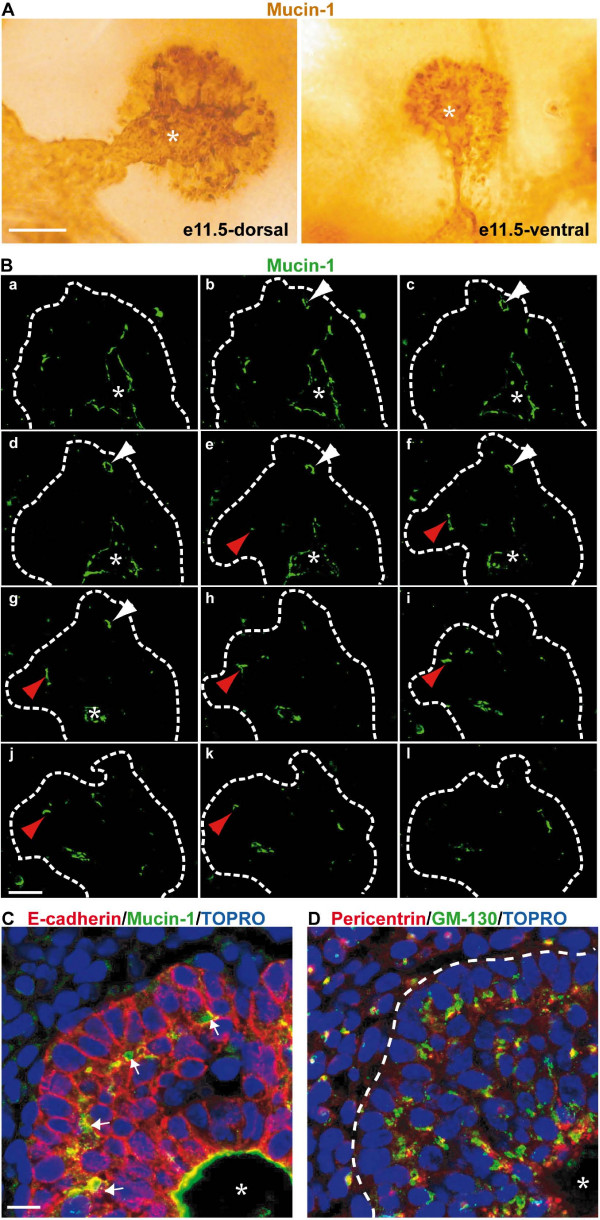
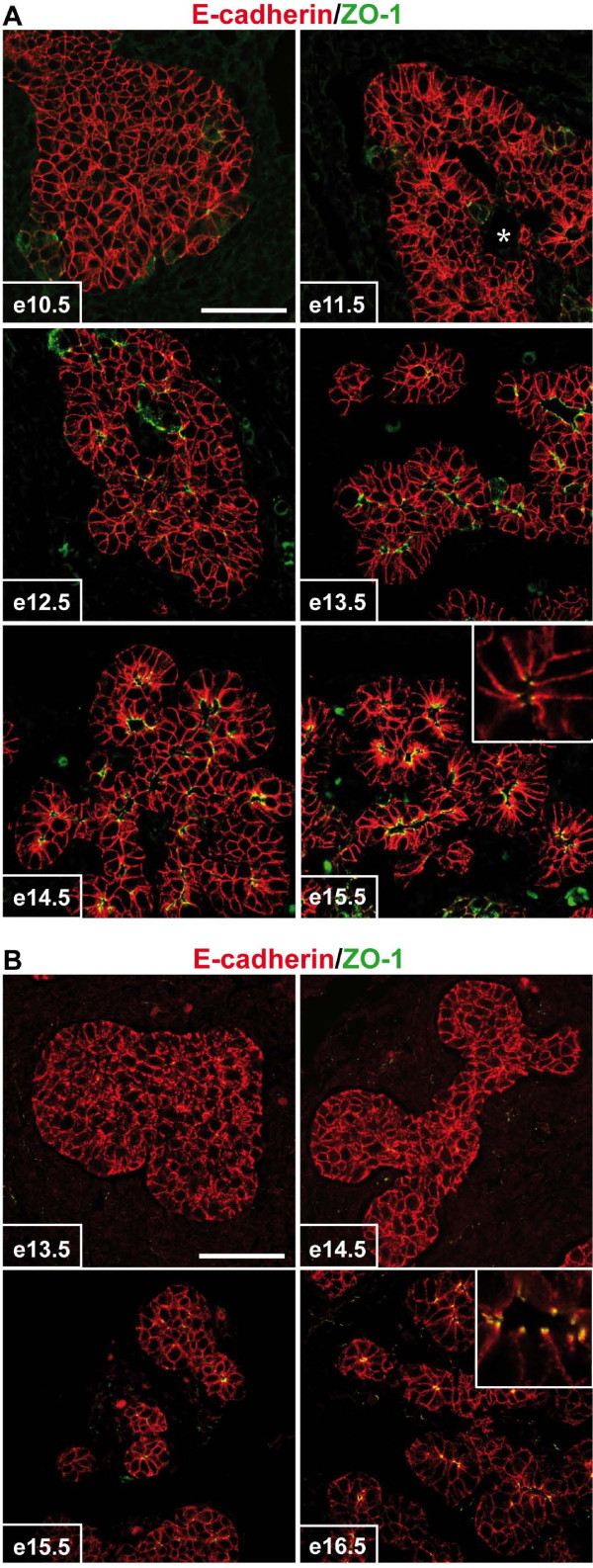
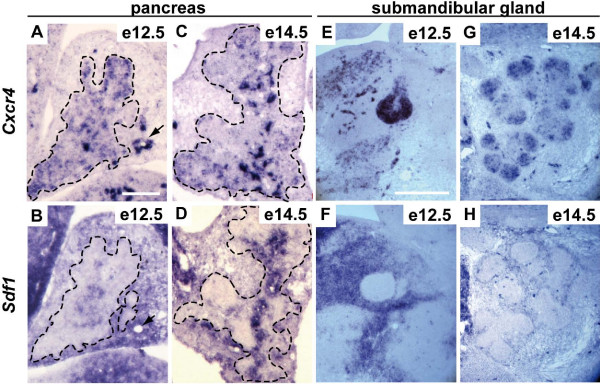
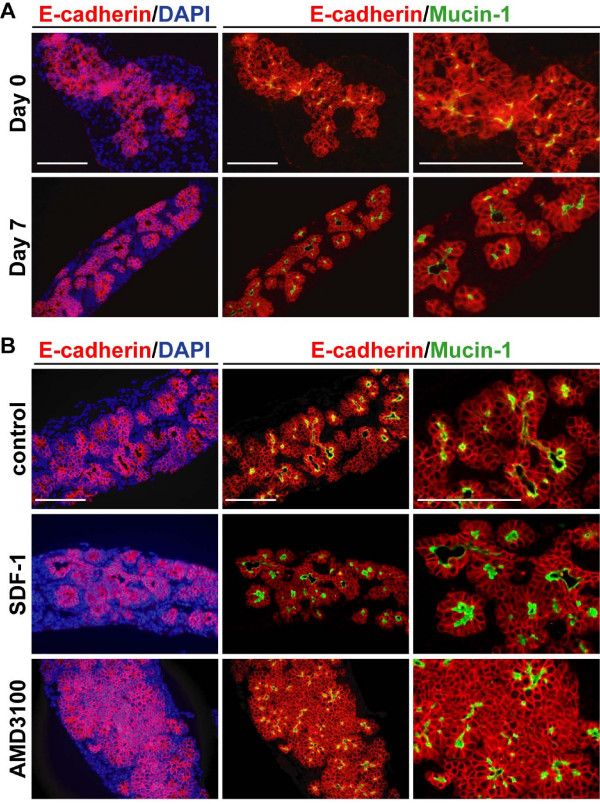
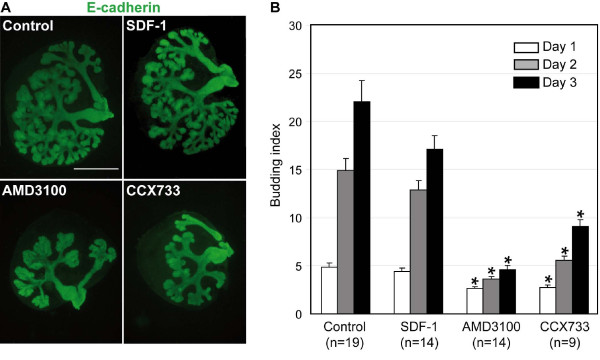
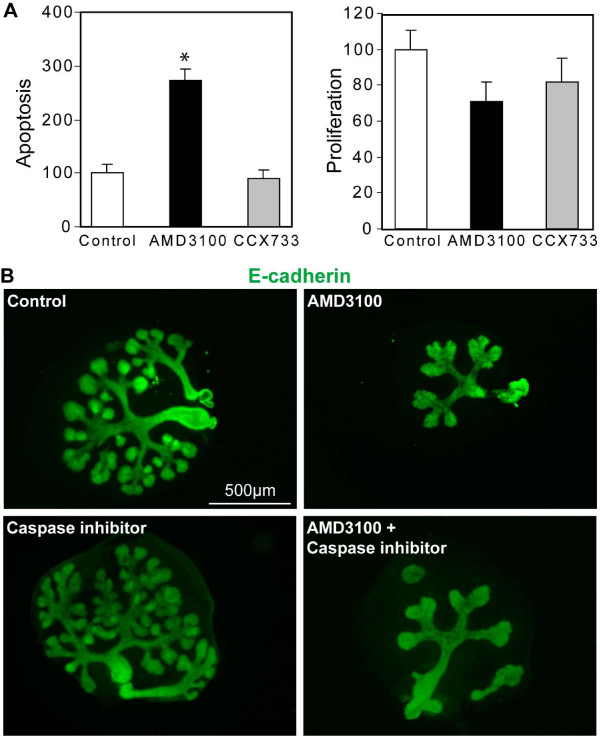
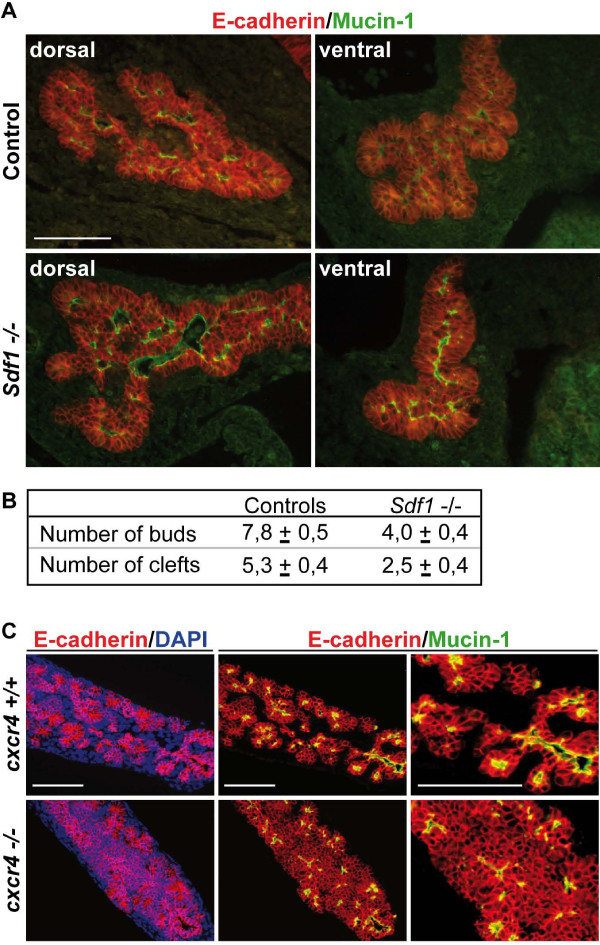
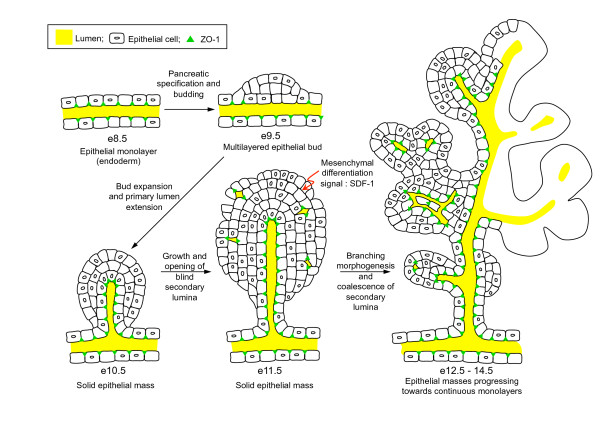
Results: By morphological analyzis of wild-type and mutant mouse embryos and using cultured embryonic explants we investigated how epithelial morphogenesis takes place and is regulated by chemokine signaling. Pancreas ontogenesis displayed a sequence of two opposite epithelial transitions. During the first transition, the monolayered and polarized endodermal cells give rise to tissue buds composed of a mass of non polarized epithelial cells. During the second transition the buds reorganize into branched and polarized epithelial monolayers that further differentiate into tubulo-acinar glands. We found that the second epithelial transition is controlled by the chemokine Stromal cell-Derived Factor (SDF)-1. The latter is expressed by the mesenchyme, whereas its receptor CXCR4 is expressed by the epithelium. Reorganization of cultured pancreatic buds into monolayered epithelia was blocked in the presence of AMD3100, a SDF-1 antagonist. Analyzis of sdf1 and cxcr4 knockout embryos at the stage of the second epithelial transition revealed transient defective morphogenesis of the ventral and dorsal pancreas. Reorganization of a globular mass of epithelial cells in polarized monolayers is also observed during submandibular glands development. We found that SDF-1 and CXCR4 are expressed in this organ and that AMD3100 treatment of submandibular gland explants blocks its branching morphogenesis.
Conclusion: In conclusion, our data show that the primitive pancreatic ductal network, which is lined by a monolayered and polarized epithelium, forms by remodeling of a globular mass of non polarized epithelial cells. Our data also suggest that SDF-1 controls the branching morphogenesis of several exocrine tissues.
Figures
References
-
- Slack JM. Developmental biology of the pancreas. Development. 1995;121:1569–1580. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
