

Immunization with the attenuated plasmidless Chlamydia trachomatis L2(25667R) strain provides partial protection in a murine model of female genitourinary tract infection
- PMID: 20004265
- PMCID: PMC2821993
- DOI: 10.1016/j.vaccine.2009.11.073
Immunization with the attenuated plasmidless Chlamydia trachomatis L2(25667R) strain provides partial protection in a murine model of female genitourinary tract infection
Abstract
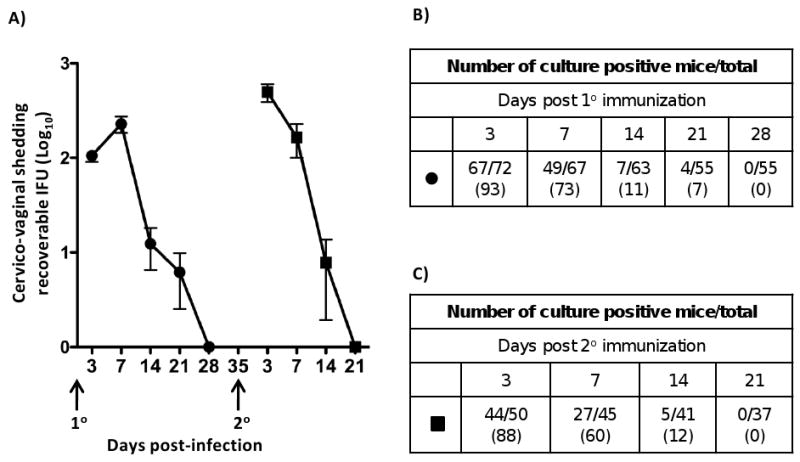
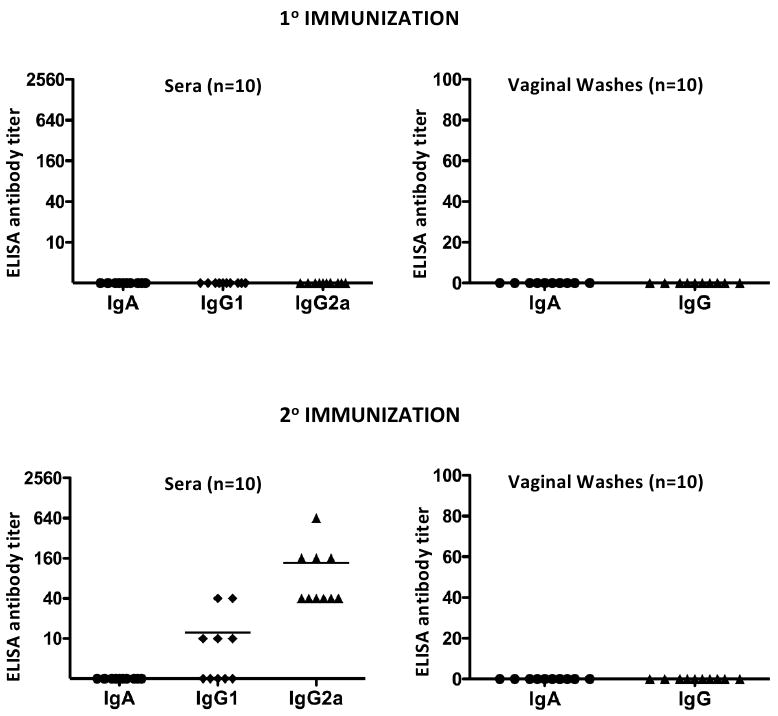
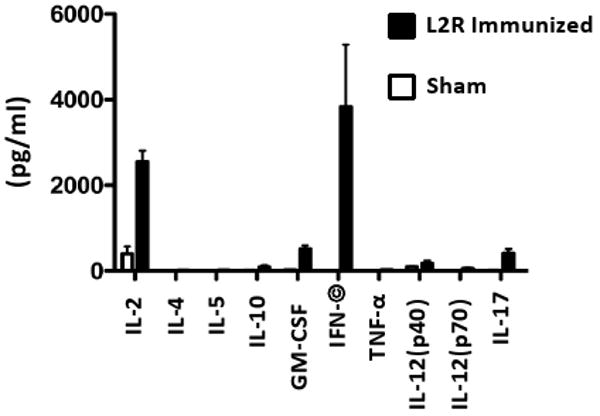
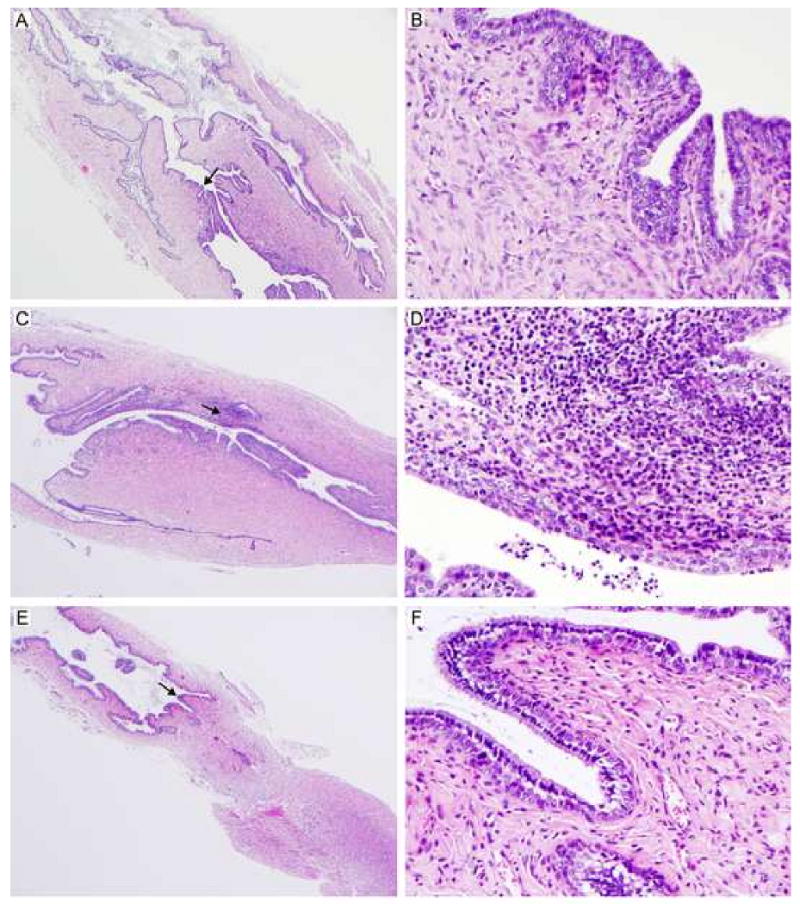
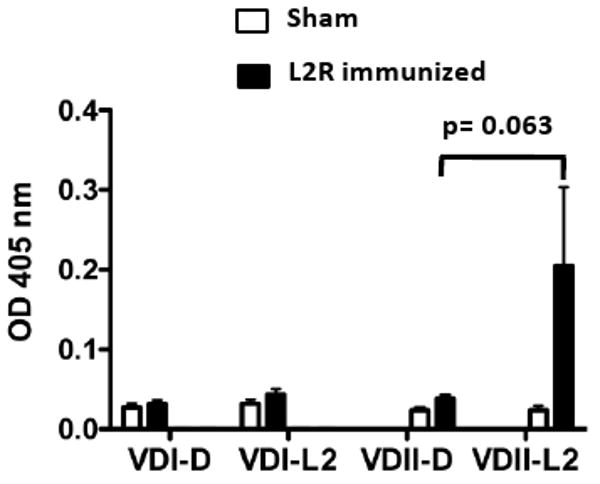
Here we report on the safety, immunogenicity, and vaccine efficacy of the naturally occurring plasmid-free attenuated Chlamydia trachomatis L2-25667R (L2R) strain in a murine infection model. Intravaginal immunization induced both chlamydial specific serum antibody and systemic CD4(+) Th1 biased immune responses but failed to induce local IgA antibodies. Immunization induced no pathological changes in the urogenital tract. Protective immunity was evaluated by vaginal challenge with a natural occurring non-attenuated plasmid positive C. trachomatis urogenital strain (serovar D). Vaccinated mice were not protected from colonization/infection but exhibited a reduction in infectious burden at early time periods (1-2 weeks) post-challenge. Partial protective immunity did not protect against inflammatory disease. Thus, intravaginal vaccination with the live-attenuated L2R stain is safe, induces a systemic antibody and CD4(+) Th1 biased immune response, but its protective efficacy is limited to reducing chlamydial burden at early time periods post-infection.
Published by Elsevier Ltd.
Figures
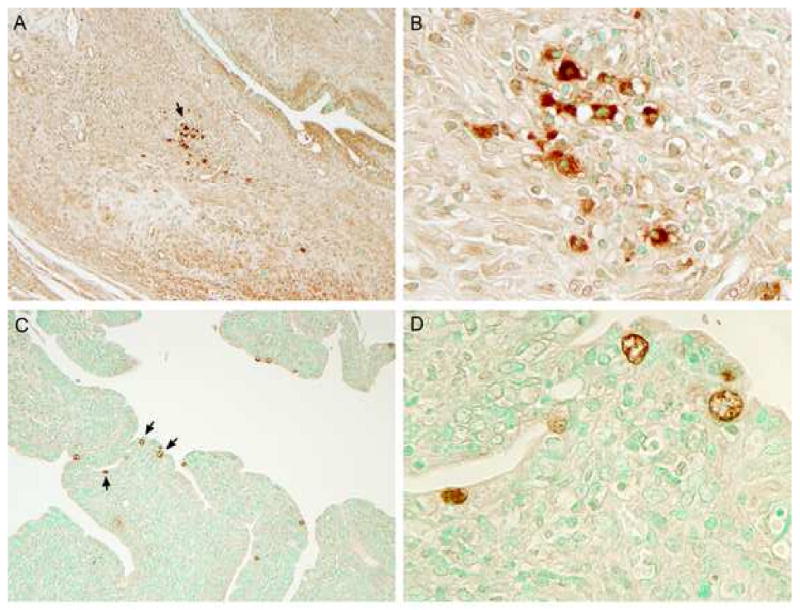
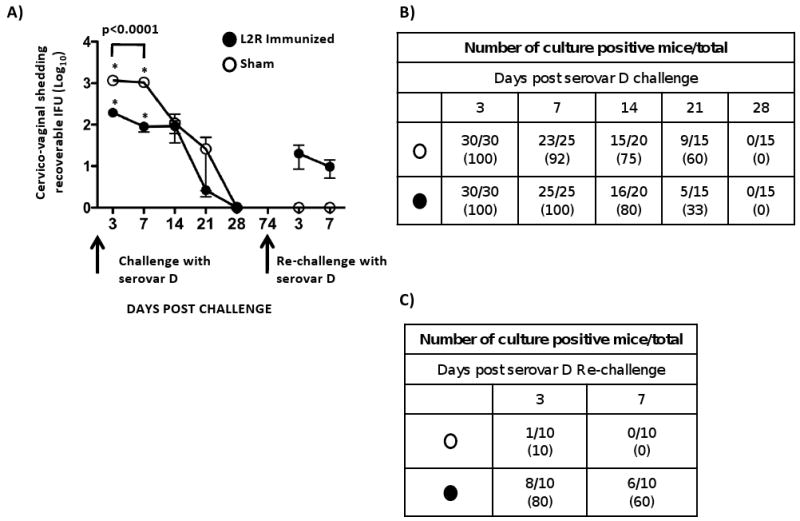
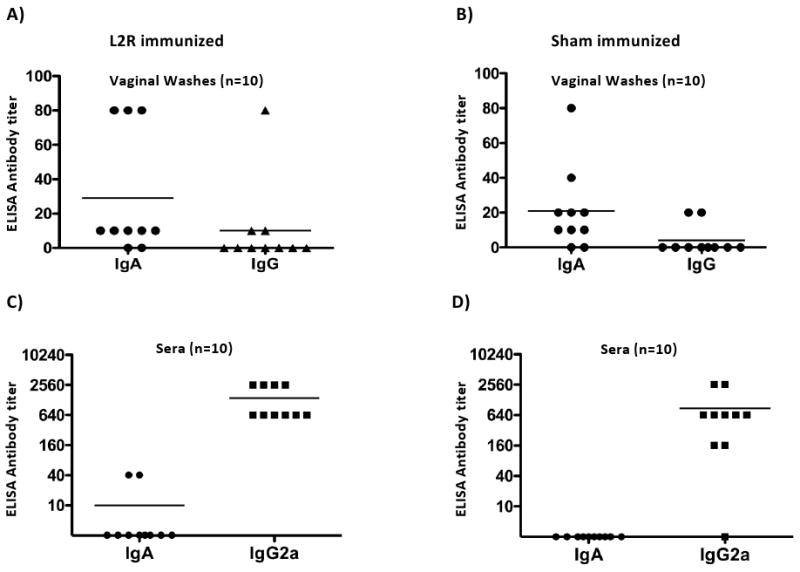
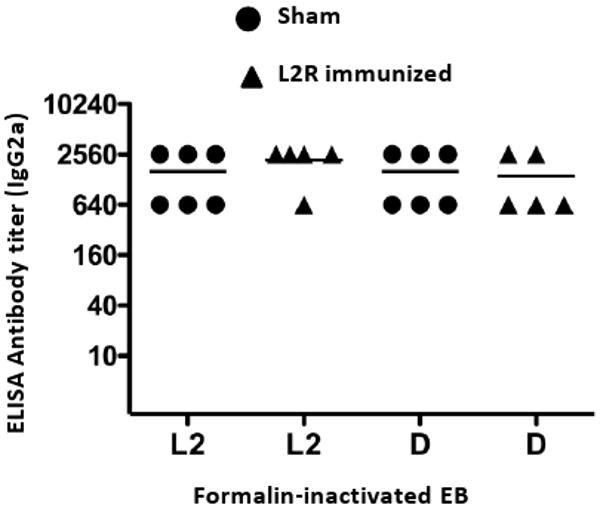
References
-
- Brunham RC, Rey-Ladino J. Immunology of Chlamydia infection: implications for a Chlamydia trachomatis vaccine. Nat Rev Immunol. 2005;(2):149–61. - PubMed
-
- Eko FO, He Q, Brown T, McMillan L, Ifere GO, Ananaba GA, et al. A novel recombinant multisubunit vaccine against Chlamydia. J Immunol. 2004;(5):3375–82. - PubMed
-
- Murthy AK, Chambers JP, Meier PA, Zhong G, Arulanandam BP. Intranasal vaccination with a secreted chlamydial protein enhances resolution of genital Chlamydia muridarum infection, protects against oviduct pathology, and is highly dependent upon endogenous gamma interferon production. Infect Immun. 2007;(2):666–76. - PMC - PubMed
-
- Su H, Parnell M, Caldwell HD. Protective efficacy of a parenterally administered MOMP-derived synthetic oligopeptide vaccine in a murine model of Chlamydia trachomatis genital tract infection: serum neutralizing IgG antibodies do not protect against chlamydial genital tract infection. Vaccine. 1995;(11):1023–32. - PubMed
-
- Dong-Ji Z, Yang X, Shen C, Lu H, Murdin A, Brunham RC. Priming with Chlamydia trachomatis major outer membrane protein (MOMP) DNA followed by MOMP ISCOM boosting enhances protection and is associated with increased immunoglobulin A and Th1 cellular immune responses. Infect Immun. 2000;(6):3074–8. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
