

Nuclear networking fashions pre-messenger RNA and primary microRNA transcripts for function
- PMID: 20004579
- PMCID: PMC2821161
- DOI: 10.1016/j.tcb.2009.10.004
Nuclear networking fashions pre-messenger RNA and primary microRNA transcripts for function
Abstract
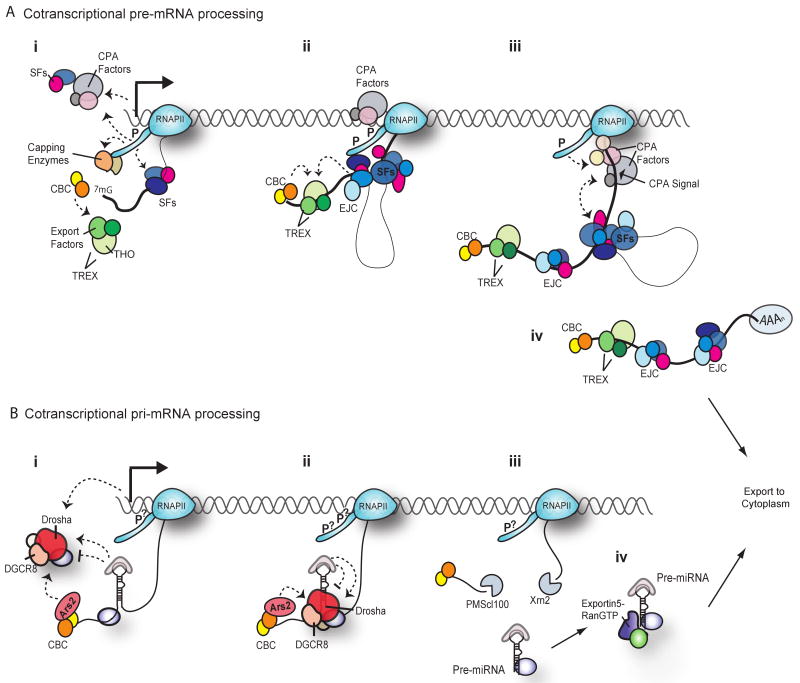
The expression of protein-coding genes is enhanced by the exquisite coupling of transcription by RNA polymerase II with pre-messenger RNA processing reactions, such as 5'-end capping, splicing and 3'-end formation. Integration between cotranscriptional processing events extends beyond the nucleus, as proteins that bind cotranscriptionally can affect the localization, translation and degradation of the mature messenger RNA. MicroRNAs are RNA polymerase II transcripts with crucial roles in the regulation of gene expression. Recent data demonstrate that processing of primary microRNA transcripts might be yet another cotranscriptional event that is woven into this elaborate nuclear network. This review discusses the extensive molecular interactions that couple the earliest steps in gene expression and therefore influence the final fate and function of the mature messenger RNA or microRNA produced.
Figures
Similar articles
-
Cellular localization and processing of primary transcripts of exonic microRNAs.PLoS One. 2013 Sep 20;8(9):e76647. doi: 10.1371/journal.pone.0076647. eCollection 2013. PLoS One. 2013. PMID: 24073292 Free PMC article.
-
Long-read sequencing of nascent RNA reveals coupling among RNA processing events.Genome Res. 2018 Jul;28(7):1008-1019. doi: 10.1101/gr.232025.117. Epub 2018 Jun 14. Genome Res. 2018. PMID: 29903723 Free PMC article.
-
Pre-mRNA splicing and its cotranscriptional connections.Trends Genet. 2023 Sep;39(9):672-685. doi: 10.1016/j.tig.2023.04.008. Epub 2023 May 24. Trends Genet. 2023. PMID: 37236814 Free PMC article. Review.
-
Balance between MAT2A intron detention and splicing is determined cotranscriptionally.RNA. 2018 Jun;24(6):778-786. doi: 10.1261/rna.064899.117. Epub 2018 Mar 21. RNA. 2018. PMID: 29563249 Free PMC article.
-
Nuclear post-transcriptional control of gene expression.J Mol Endocrinol. 2001 Oct;27(2):123-31. doi: 10.1677/jme.0.0270123. J Mol Endocrinol. 2001. PMID: 11564598 Review.
Cited by
-
RNA Pol II Assembly Affects ncRNA Expression.Int J Mol Sci. 2023 Dec 29;25(1):507. doi: 10.3390/ijms25010507. Int J Mol Sci. 2023. PMID: 38203678 Free PMC article.
-
CDKF;1 and CDKD protein kinases regulate phosphorylation of serine residues in the C-terminal domain of Arabidopsis RNA polymerase II.Plant Cell. 2012 Apr;24(4):1626-42. doi: 10.1105/tpc.112.096834. Epub 2012 Apr 30. Plant Cell. 2012. PMID: 22547781 Free PMC article.
-
Hepatic miRNA expression reprogrammed by Plasmodium chabaudi malaria.Parasitol Res. 2011 May;108(5):1111-21. doi: 10.1007/s00436-010-2152-z. Epub 2010 Nov 18. Parasitol Res. 2011. PMID: 21085987
-
Ars2 promotes proper replication-dependent histone mRNA 3' end formation.Mol Cell. 2012 Jan 13;45(1):87-98. doi: 10.1016/j.molcel.2011.12.020. Mol Cell. 2012. PMID: 22244333 Free PMC article.
-
Circadian control of mRNA polyadenylation dynamics regulates rhythmic protein expression.Genes Dev. 2012 Dec 15;26(24):2724-36. doi: 10.1101/gad.208306.112. Genes Dev. 2012. PMID: 23249735 Free PMC article.
References
-
- Neugebauer KM. On the importance of being co-transcriptional. J Cell Sci. 2002;115:3865–3871. - PubMed
-
- Bentley DL. Rules of engagement: co-transcriptional recruitment of pre-mRNA processing factors. Curr Opin Cell Biol. 2005;17:251–256. - PubMed
-
- Matera AG, et al. Non-coding RNAs: lessons from the small nuclear and small nucleolar RNAs. Nat Rev Mol Cell Biol. 2007;8:209–220. - PubMed
-
- Gregory RI, et al. The Microprocessor complex mediates the genesis of microRNAs. Nature. 2004;432:235–240. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
