

Preferential ethanol consumption in Drosophila models features of addiction
- PMID: 20005106
- PMCID: PMC2805771
- DOI: 10.1016/j.cub.2009.10.070
Preferential ethanol consumption in Drosophila models features of addiction
Abstract
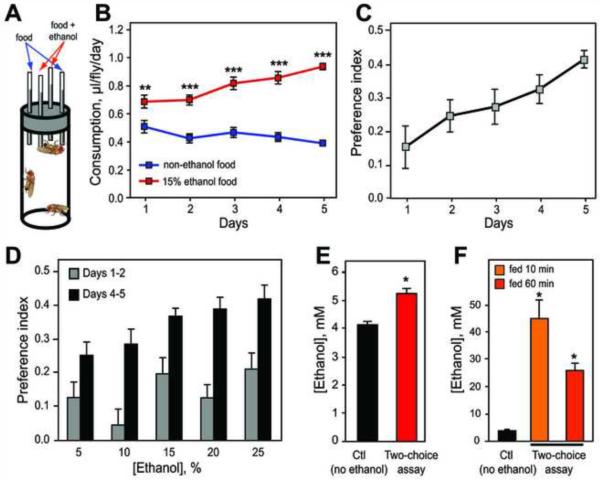
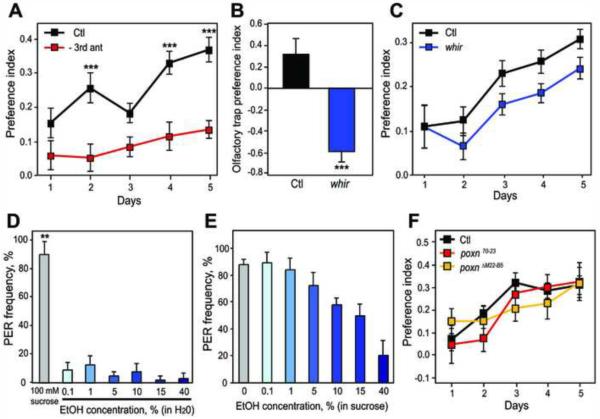
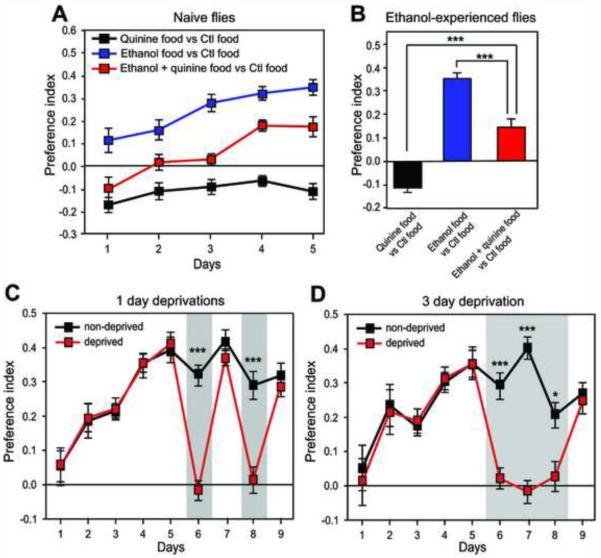
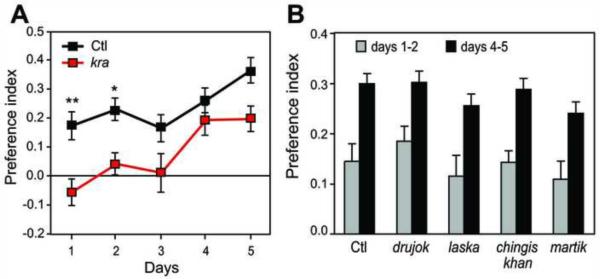
Alcohol addiction is a common affliction with a strong genetic component [1]. Although mammalian studies have provided significant insight into the molecular mechanisms underlying ethanol consumption [2], other organisms such as Drosophila melanogaster are better suited for unbiased, forward genetic approaches to identify novel genes. Behavioral responses to ethanol, such as hyperactivity, sedation, and tolerance, are conserved between flies and mammals [3, 4], as are the underlying molecular pathways [5-9]. However, few studies have investigated ethanol self-administration in flies [10]. Here we characterize ethanol consumption and preference in Drosophila. Flies prefer to consume ethanol-containing food over regular food, and this preference increases over time. Flies are attracted to the smell of ethanol, which partially mediates ethanol preference, but are averse to its taste. Preference for consuming ethanol is not entirely explained by attraction to either its sensory or caloric properties. We demonstrate that flies can exhibit features of alcohol addiction. First, flies self-administer ethanol to pharmacologically relevant concentrations. Second, flies will overcome an aversive stimulus in order to consume ethanol. Third, flies rapidly return to high levels of ethanol consumption after a period of imposed abstinence. Thus, ethanol preference in Drosophila provides a new model for studying aspects of addiction.
Figures
References
-
- Dick DM, Foroud T. Candidate genes for alcohol dependence: a review of genetic evidence from human studies. Alcohol Clin. Exp. Res. 2003;27:868–879. - PubMed
-
- Crabbe JC, Phillips TJ, Harris RA, Arends MA, Koob GF. Alcohol-related genes: contributions from studies with genetically engineered mice. Addict. Biol. 2006;11:195–269. - PubMed
-
- Scholz H, Ramond J, Singh CM, Heberlein U. Functional ethanol tolerance in Drosophila. Neuron. 2000;28:261–271. - PubMed
-
- Moore MS, DeZazzo J, Luk AY, Tully T, Singh CM, Heberlein U. Ethanol intoxication in Drosophila: genetic and pharmacological evidence for regulation by the cAMP signaling pathway. Cell. 1998;93:997–1007. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
