

The structural basis for mRNA recognition and cleavage by the ribosome-dependent endonuclease RelE
- PMID: 20005802
- PMCID: PMC2807027
- DOI: 10.1016/j.cell.2009.11.015
The structural basis for mRNA recognition and cleavage by the ribosome-dependent endonuclease RelE
Abstract
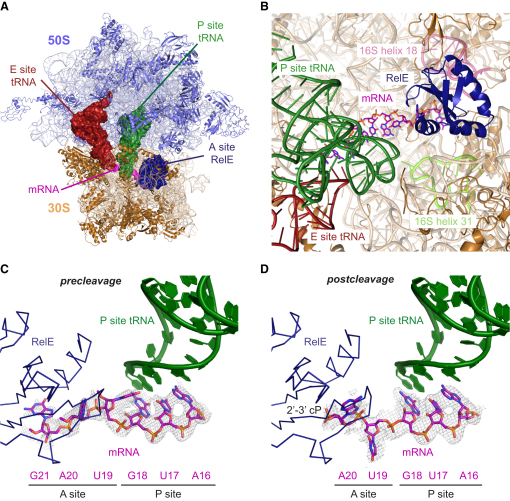
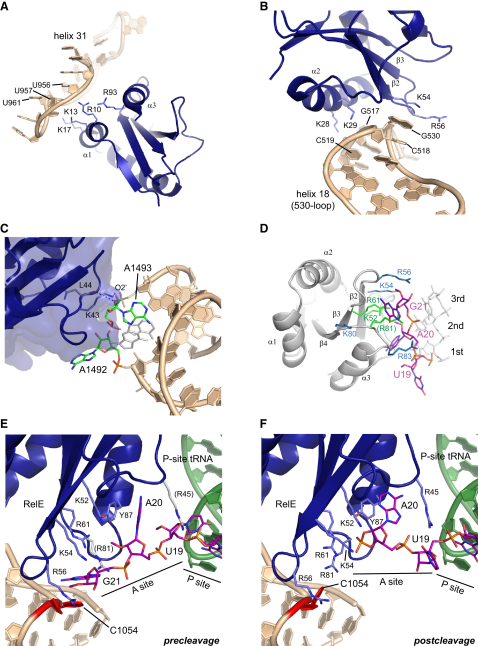
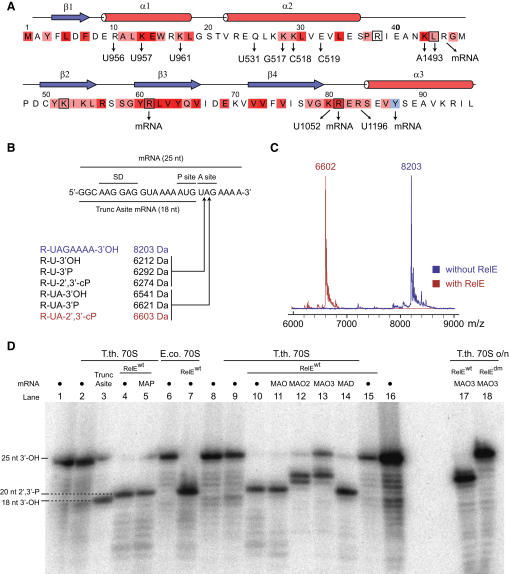
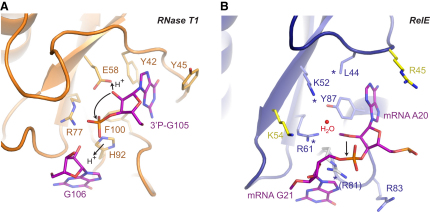
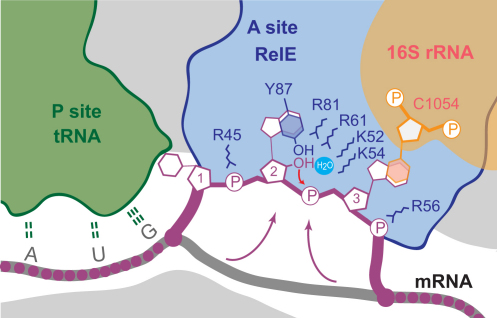
Translational control is widely used to adjust gene expression levels. During the stringent response in bacteria, mRNA is degraded on the ribosome by the ribosome-dependent endonuclease, RelE. The molecular basis for recognition of the ribosome and mRNA by RelE and the mechanism of cleavage are unknown. Here, we present crystal structures of E. coli RelE in isolation (2.5 A) and bound to programmed Thermus thermophilus 70S ribosomes before (3.3 A) and after (3.6 A) cleavage. RelE occupies the A site and causes cleavage of mRNA after the second nucleotide of the codon by reorienting and activating the mRNA for 2'-OH-induced hydrolysis. Stacking of A site codon bases with conserved residues in RelE and 16S rRNA explains the requirement for the ribosome in catalysis and the subtle sequence specificity of the reaction. These structures provide detailed insight into the translational regulation on the bacterial ribosome by mRNA cleavage.
Figures

Similar articles
-
Molecular basis of ribosome recognition and mRNA hydrolysis by the E. coli YafQ toxin.Nucleic Acids Res. 2015 Sep 18;43(16):8002-12. doi: 10.1093/nar/gkv791. Epub 2015 Aug 10. Nucleic Acids Res. 2015. PMID: 26261214 Free PMC article.
-
A ribosome profiling study of mRNA cleavage by the endonuclease RelE.Nucleic Acids Res. 2017 Jan 9;45(1):327-336. doi: 10.1093/nar/gkw944. Epub 2016 Oct 18. Nucleic Acids Res. 2017. PMID: 27924019 Free PMC article.
-
Transition State Charge Stabilization and Acid-Base Catalysis of mRNA Cleavage by the Endoribonuclease RelE.Biochemistry. 2015 Dec 1;54(47):7048-57. doi: 10.1021/acs.biochem.5b00866. Epub 2015 Nov 12. Biochemistry. 2015. PMID: 26535789 Free PMC article.
-
The bacterial toxin RelE induces specific mRNA cleavage in the A site of the eukaryote ribosome.RNA. 2008 Feb;14(2):233-9. doi: 10.1261/rna.693208. Epub 2007 Dec 14. RNA. 2008. PMID: 18083838 Free PMC article.
-
Ribosome structure and dynamics during translocation and termination.Annu Rev Biophys. 2010;39:227-44. doi: 10.1146/annurev.biophys.37.032807.125954. Annu Rev Biophys. 2010. PMID: 20192776 Review.
Cited by
-
The higBA-Type Toxin-Antitoxin System in IncC Plasmids Is a Mobilizable Ciprofloxacin-Inducible System.mSphere. 2021 Jun 30;6(3):e0042421. doi: 10.1128/mSphere.00424-21. Epub 2021 Jun 2. mSphere. 2021. PMID: 34077263 Free PMC article.
-
Solution structure and membrane binding of the toxin fst of the par addiction module.Biochemistry. 2010 Aug 10;49(31):6567-75. doi: 10.1021/bi1005128. Biochemistry. 2010. PMID: 20677831 Free PMC article.
-
The relBE2Spn toxin-antitoxin system of Streptococcus pneumoniae: role in antibiotic tolerance and functional conservation in clinical isolates.PLoS One. 2010 Jun 23;5(6):e11289. doi: 10.1371/journal.pone.0011289. PLoS One. 2010. PMID: 20585658 Free PMC article.
-
Regulating toxin-antitoxin expression: controlled detonation of intracellular molecular timebombs.Toxins (Basel). 2014 Jan 15;6(1):337-58. doi: 10.3390/toxins6010337. Toxins (Basel). 2014. PMID: 24434949 Free PMC article. Review.
-
Enteric virulence associated protein VapC inhibits translation by cleavage of initiator tRNA.Proc Natl Acad Sci U S A. 2011 May 3;108(18):7403-7. doi: 10.1073/pnas.1019587108. Epub 2011 Apr 18. Proc Natl Acad Sci U S A. 2011. PMID: 21502523 Free PMC article.
References
-
- Adams P.D., Grosse-Kunstleve R.W., Hung L.W., Ioerger T.R., McCoy A.J., Moriarty N.W., Read R.J., Sacchettini J.C., Sauter N.K., Terwilliger T.C. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr. D Biol. Crystallogr. 2002;58:1948–1954. - PubMed
-
- Bauerova-Hlinkova V., Dvorsky R., Perecko D., Povazanec F., Sevcik J. Structure of RNase Sa2 complexes with mononucleotides–new aspects of catalytic reaction and substrate recognition. FEBS J. 2009;276:4156–4168. - PubMed
-
- Brunger A.T., Adams P.D., Clore G.M., DeLano W.L., Gros P., Grosse-Kunstleve R.W., Jiang J.S., Kuszewski J., Nilges M., Pannu N.S. Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr. D Biol. Crystallogr. 1998;54:905–921. - PubMed
-
- Christensen-Dalsgaard M., Gerdes K. Two higBA loci in the Vibrio cholerae superintegron encode mRNA cleaving enzymes and can stabilize plasmids. Mol. Microbiol. 2006;62:397–411. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
