

Rrp17p is a eukaryotic exonuclease required for 5' end processing of Pre-60S ribosomal RNA
- PMID: 20005841
- PMCID: PMC2806520
- DOI: 10.1016/j.molcel.2009.11.011
Rrp17p is a eukaryotic exonuclease required for 5' end processing of Pre-60S ribosomal RNA
Abstract
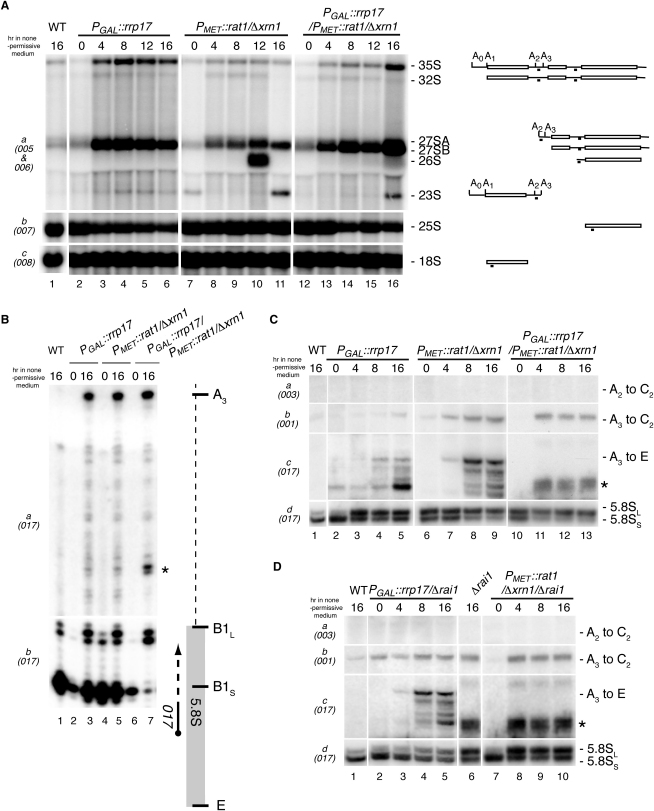
Ribosomal processing requires a series of endo- and exonucleolytic steps for the production of mature ribosomes, of which most have been described. To ensure ribosome synthesis, 3' end formation of rRNA uses multiple nucleases acting in parallel; however, a similar parallel mechanism had not been described for 5' end maturation. Here, we identify Rrp17p as a previously unidentified 5'-3' exonuclease essential for ribosome biogenesis, functioning with Rat1p in a parallel processing pathway analogous to that of 3' end formation. Rrp17p is required for efficient exonuclease digestion of the mature 5' ends of 5.8S(S) and 25S rRNAs, contains a catalytic domain close to its N terminus, and is highly conserved among higher eukaryotes, being a member of a family of exonucleases. We show that Rrp17p binds late pre-60S ribosomes, accompanying them from the nucleolus to the nuclear periphery, and provide evidence for physical and functional links between late 60S subunit processing and export.
Figures






References
-
- Alber F., Dokudovskaya S., Veenhoff L.M., Zhang W., Kipper J., Devos D., Suprapto A., Karni-Schmidt O., Williams R., Chait B.T. Determining the architectures of macromolecular assemblies. Nature. 2007;450:683–694. - PubMed
-
- Bassler J., Grandi P., Gadal O., Lessmann T., Petfalski E., Tollervey D., Lechner J., Hurt E. Identification of a 60S preribosomal particle that is closely linked to nuclear export. Mol. Cell. 2001;8:517–529. - PubMed
-
- Briggs M.W., Burkard K.T., Butler J.S. Rrp6p, the yeast homologue of the human PM-Scl 100-kDa autoantigen, is essential for efficient 5.8 S rRNA 3′ end formation. J. Biol. Chem. 1998;273:13255–13263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
