

Ubiquitinated proteins activate the proteasome by binding to Usp14/Ubp6, which causes 20S gate opening
- PMID: 20005843
- PMCID: PMC2796264
- DOI: 10.1016/j.molcel.2009.11.015
Ubiquitinated proteins activate the proteasome by binding to Usp14/Ubp6, which causes 20S gate opening
Abstract
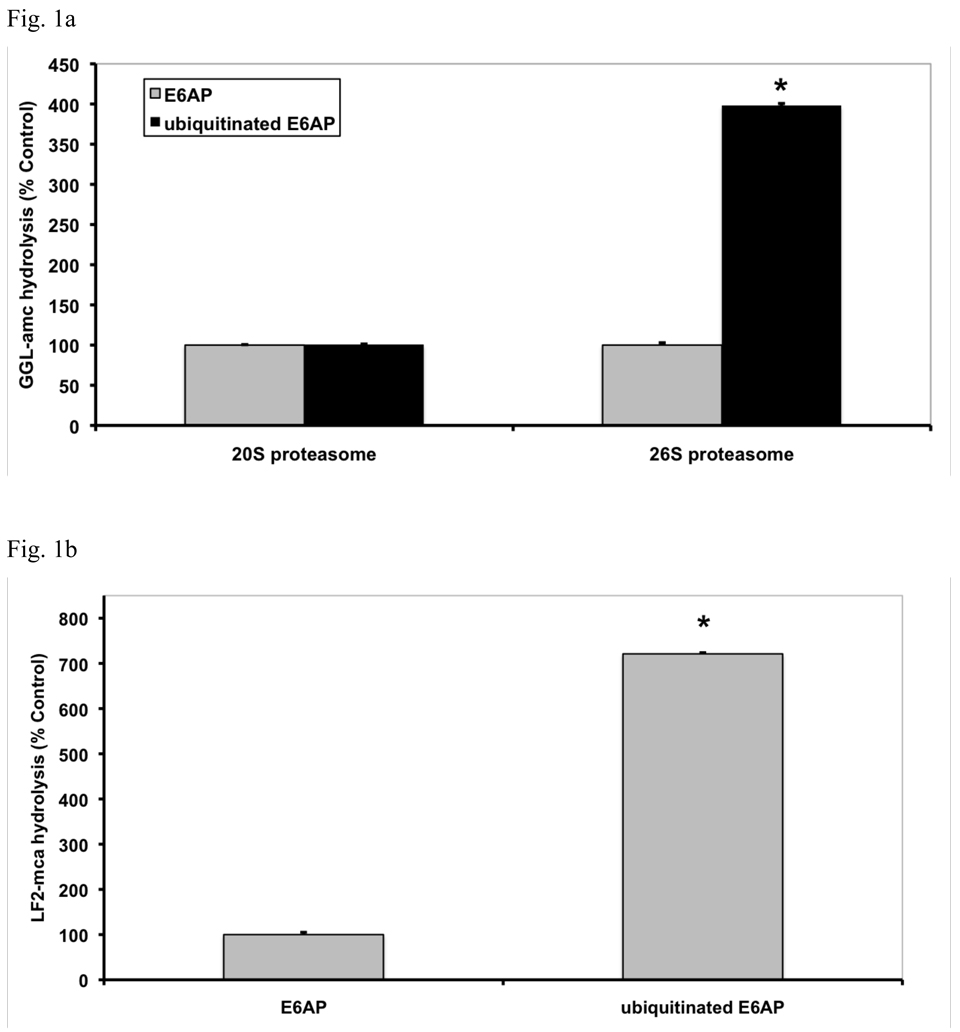
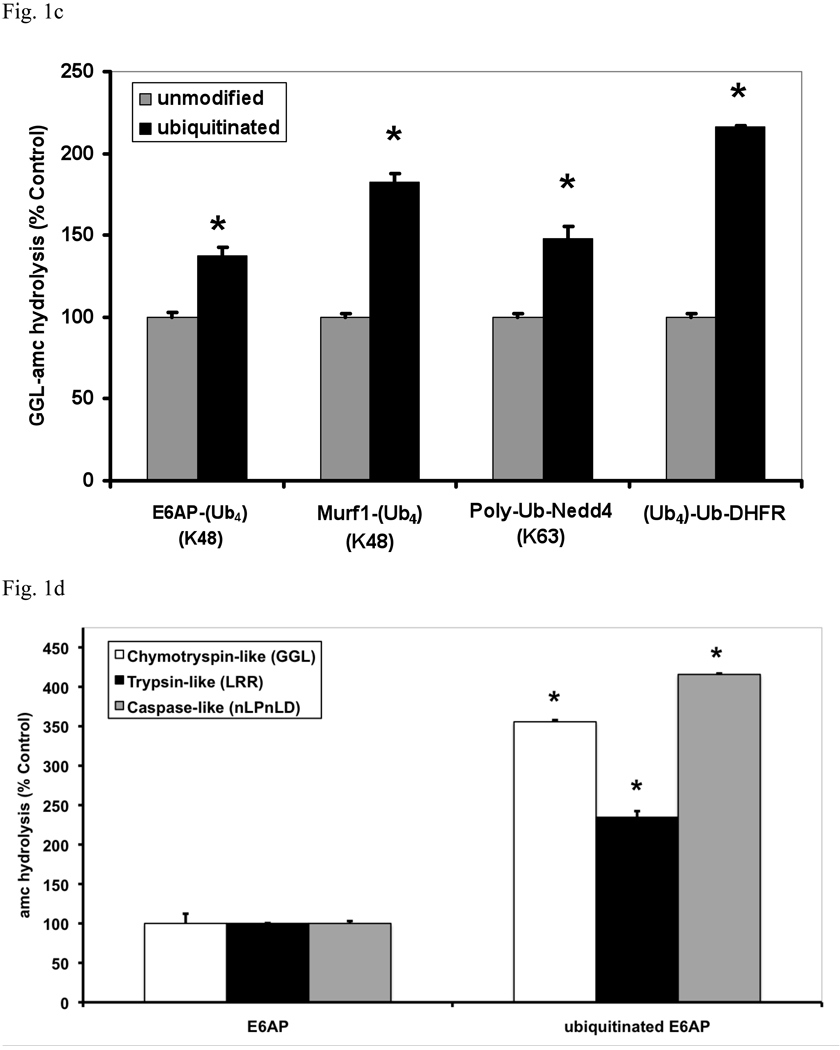
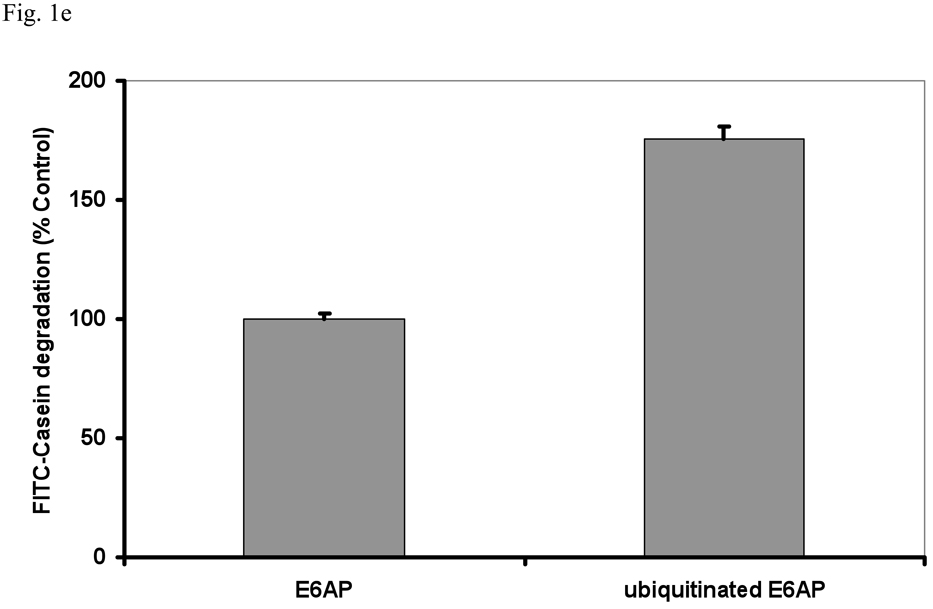
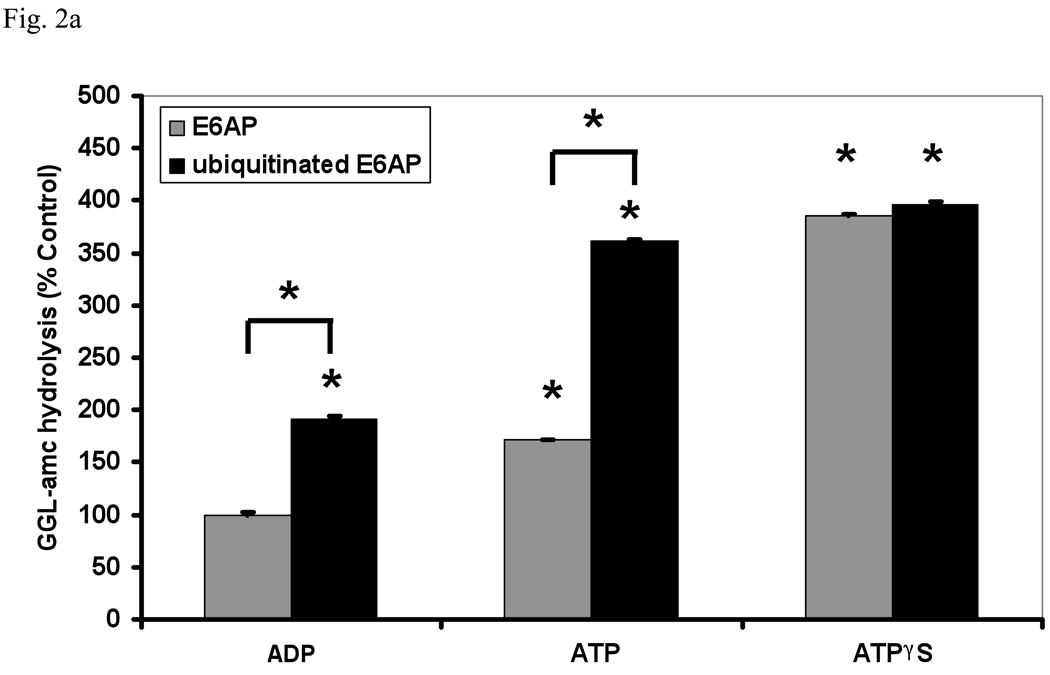
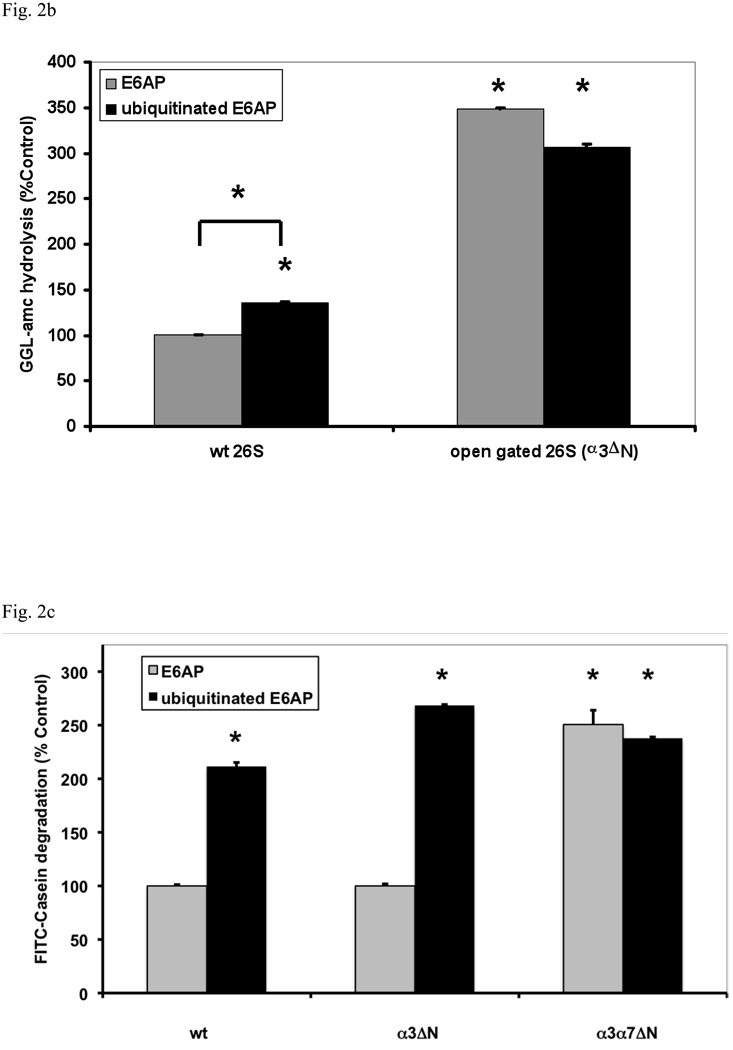
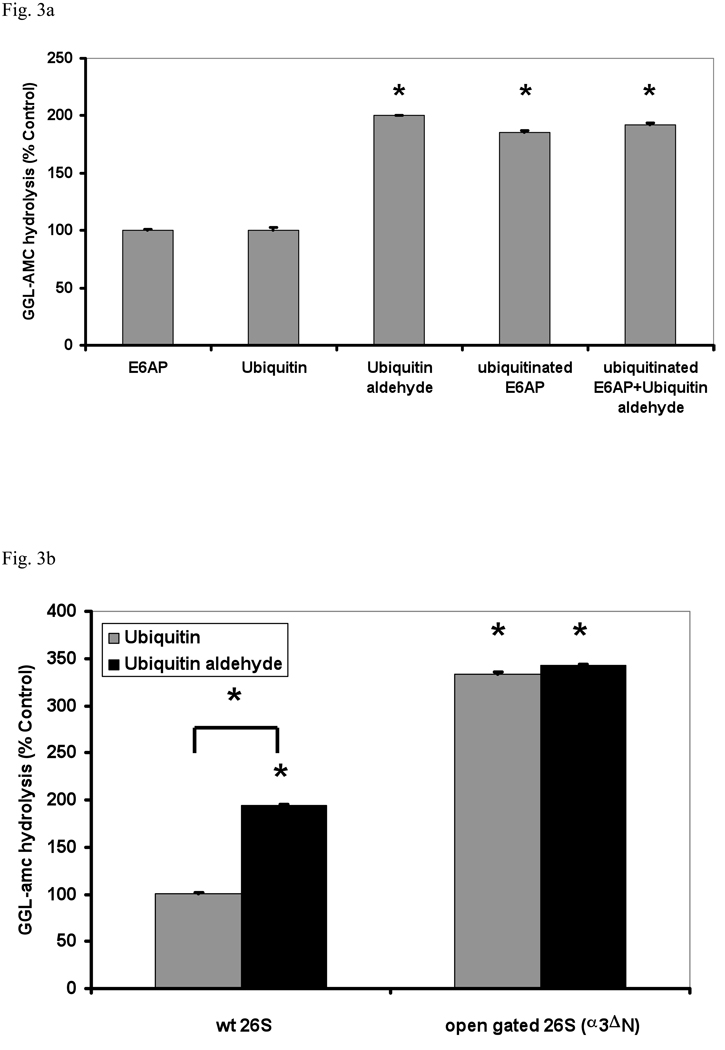
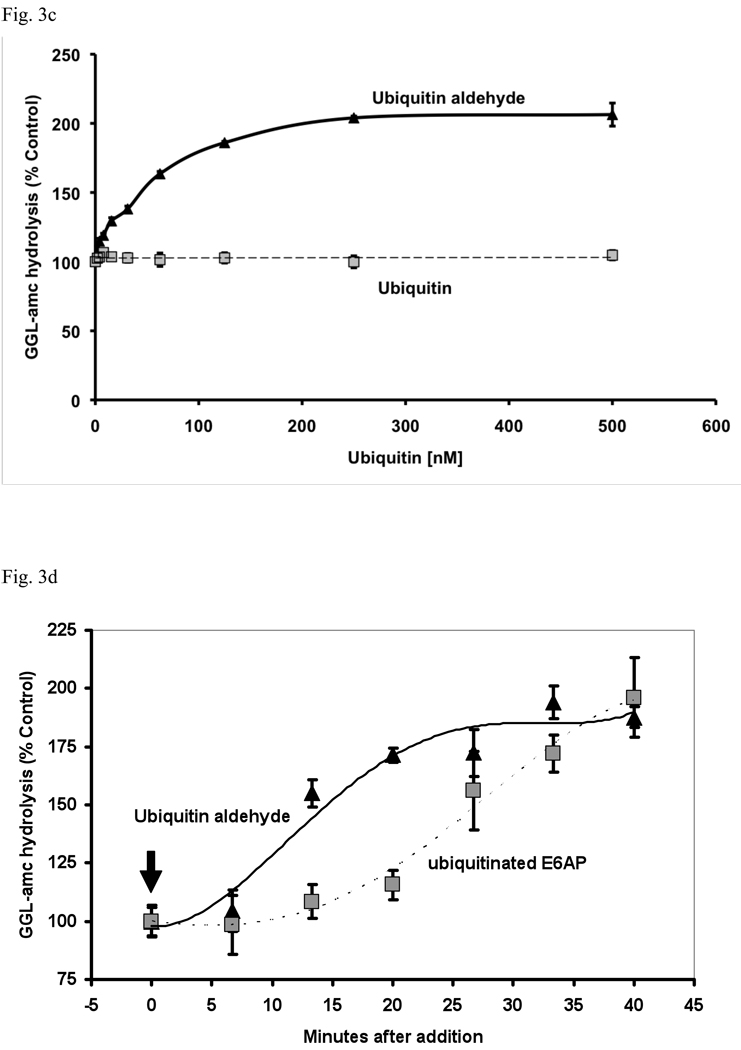
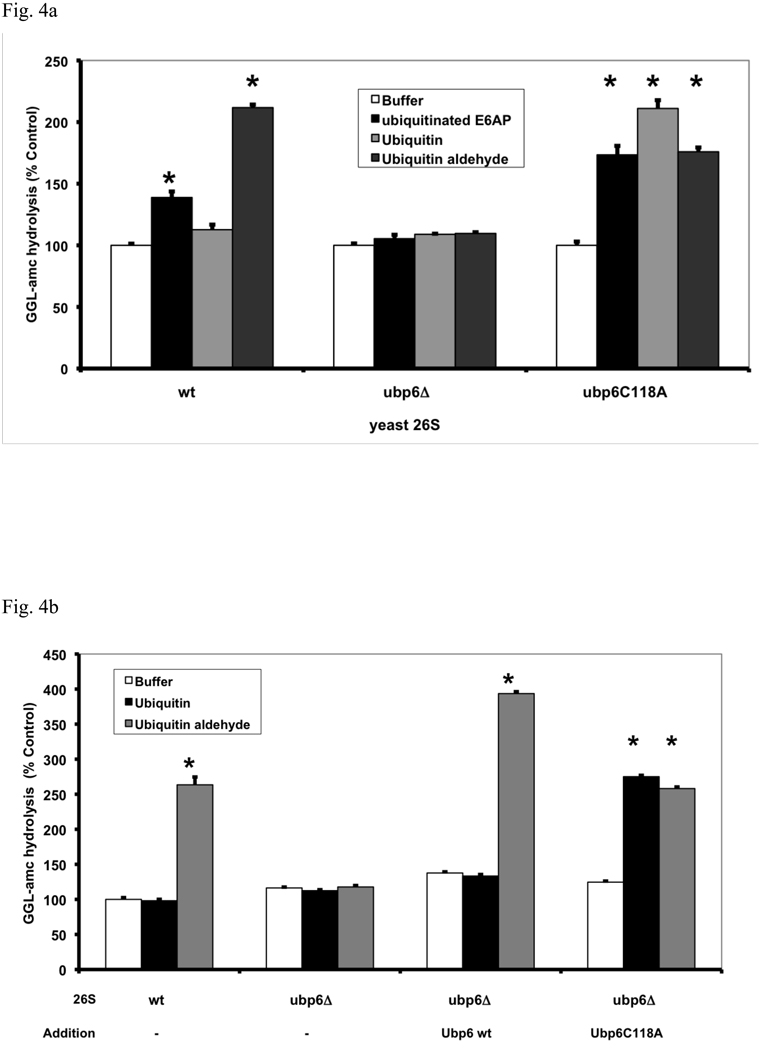
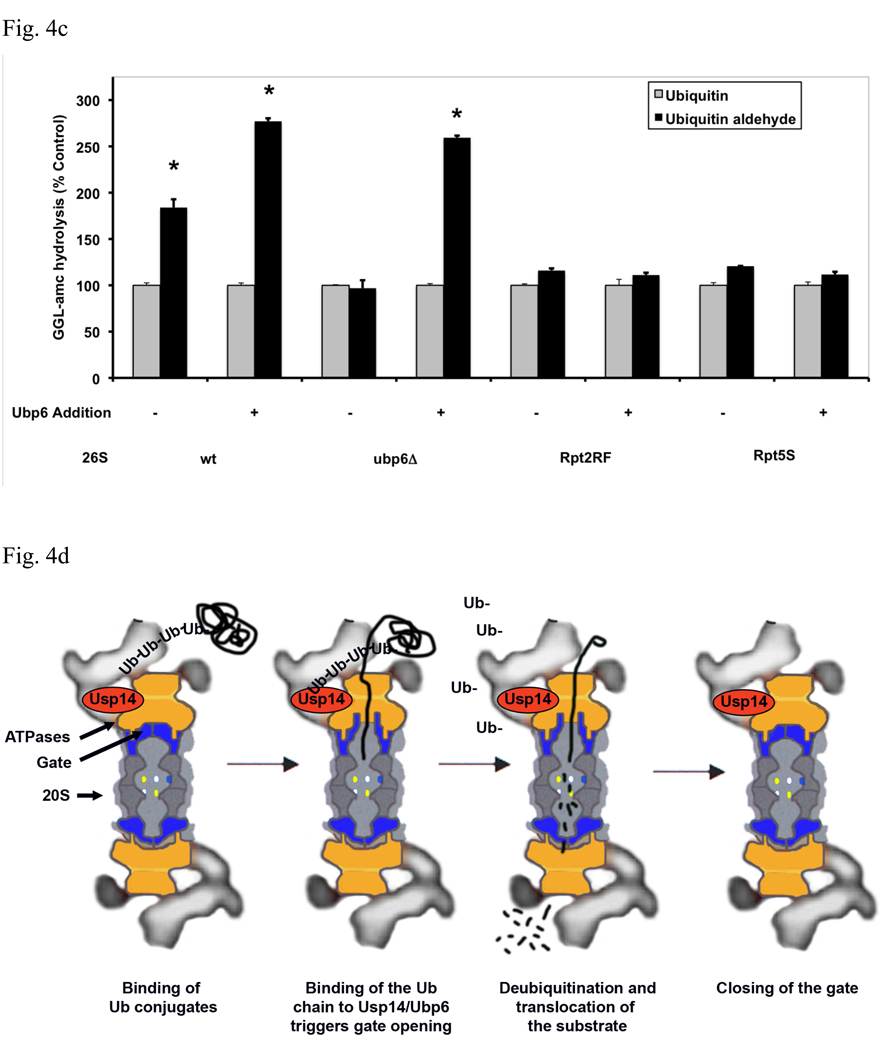
In eukaryotic cells, ubiquitination of proteins leads to their degradation by the 26S proteasome. We tested if the ubiquitin (Ub) chain also regulates the proteasome's capacity for proteolysis. After incubation with polyubiquitinated proteins, 26S proteasomes hydrolyzed peptides and proteins 2- to 7-fold faster. Ub conjugates enhanced peptide hydrolysis by stimulating gate opening in the 20S proteasome. This stimulation was seen when this gate was closed or transiently open, but not maximally open. Gate opening requires conjugate association with Usp14/Ubp6 and also occurs if Ub aldehyde occupies this isopeptidase's active site. No stimulation was observed with 26S from Ubp6Delta mutants, but this effect was restored upon addition of Usp14/Ubp6 (even an inactive Ubp6). The stimulation of gate opening by Ub conjugates through Usp14/Ubp6 requires nucleotide binding to the gate-regulatory ATPases. This activation enhances the selectivity of the 26S proteasome for ubiquitinated proteins and links their deubiquitination to their degradation.
Figures
References
-
- Bajorek M, Finley D, Glickman MH. Proteasome disassembly and downregulation is correlated with viability during stationary phase. Curr Biol. 2003;13:1140–1144. - PubMed
-
- Bech-Otschir D, Helfrich A, Enenkel C, Consiglieri G, Seeger M, Holzhutter HG, Dahlmann B, Kloetzel PM. Polyubiquitin substrates allosterically activate their own degradation by the 26S proteasome. Nat Struct Mol Biol. 2009;16:219–225. - PubMed
-
- Benaroudj N, Zwickl P, Seemuller E, Baumeister W, Goldberg AL. ATP hydrolysis by the proteasome regulatory complex PAN serves multiple functions in protein degradation. Mol Cell. 2003;11:69–78. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
