

Z proteins of New World arenaviruses bind RIG-I and interfere with type I interferon induction
- PMID: 20007272
- PMCID: PMC2812374
- DOI: 10.1128/JVI.01362-09
Z proteins of New World arenaviruses bind RIG-I and interfere with type I interferon induction
Abstract
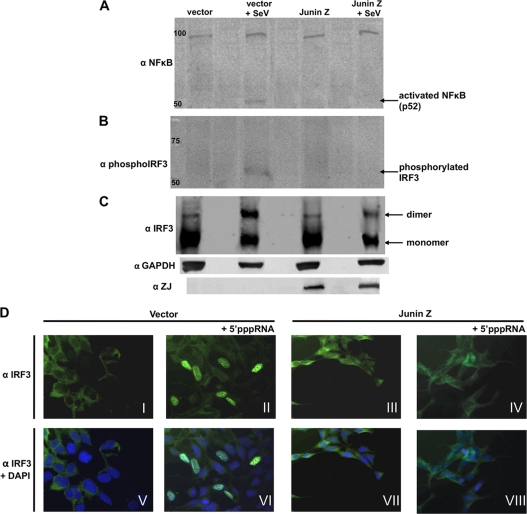
The retinoic acid-inducible gene I product (RIG-I) is a cellular sensor of RNA virus infection that regulates the cellular beta interferon (IFN-beta) response. The nucleoproteins (NP) of arenaviruses are reported to antagonize the IFN response by inhibiting interferon regulatory factor 3 (IRF-3). Here, we demonstrate that the Z proteins of four New World (NW) arenaviruses, Guanarito virus (GTOV), Junin virus (JUNV), Machupo virus (MAVC), and Sabia virus (SABV), bind to RIG-I, resulting in downregulation of the IFN-beta response. We show that expression of the four NW arenavirus Z proteins inhibits IFN-beta mRNA induction in A549 cells in response to RNA bearing 5' phosphates (5'pppRNA). NW arenavirus Z proteins interact with RIG-I in coimmunoprecipitation studies and colocalize with RIG-I. Furthermore, expression of Z proteins interferes with the interaction between RIG-I and MAVS. Z expression also impedes the nuclear factor kappa light chain enhancer of activated B cells (NF-kappaB) and IRF-3 activation. Our results indicate that NW arenavirus Z proteins, but not Z protein of the Old World (OW) arenavirus lymphocytic choriomeningitis virus (LCMV) or Lassa virus, bind to RIG-I and inhibit downstream activation of the RIG-I signaling pathway, preventing the transcriptional induction of IFN-beta.
Figures




Similar articles
-
The Z proteins of pathogenic but not nonpathogenic arenaviruses inhibit RIG-I-like receptor-dependent interferon production.J Virol. 2015 Mar;89(5):2944-55. doi: 10.1128/JVI.03349-14. Epub 2014 Dec 31. J Virol. 2015. PMID: 25552708 Free PMC article.
-
Highly Pathogenic New World Arenavirus Infection Activates the Pattern Recognition Receptor Protein Kinase R without Attenuating Virus Replication in Human Cells.J Virol. 2017 Sep 27;91(20):e01090-17. doi: 10.1128/JVI.01090-17. Print 2017 Oct 15. J Virol. 2017. PMID: 28794024 Free PMC article.
-
Lymphocytic Choriomeningitis Virus Differentially Affects the Virus-Induced Type I Interferon Response and Mitochondrial Apoptosis Mediated by RIG-I/MAVS.J Virol. 2015 Jun;89(12):6240-50. doi: 10.1128/JVI.00610-15. Epub 2015 Apr 1. J Virol. 2015. PMID: 25833049 Free PMC article.
-
Innate immune response to arenaviral infection: a focus on the highly pathogenic New World hemorrhagic arenaviruses.J Mol Biol. 2013 Dec 13;425(24):4893-903. doi: 10.1016/j.jmb.2013.09.028. Epub 2013 Sep 26. J Mol Biol. 2013. PMID: 24075870 Free PMC article. Review.
-
The catcher in the RIG-I.Cytokine. 2015 Nov;76(1):38-41. doi: 10.1016/j.cyto.2015.07.002. Epub 2015 Jul 10. Cytokine. 2015. PMID: 26168692 Review.
Cited by
-
Junín virus infection activates the type I interferon pathway in a RIG-I-dependent manner.PLoS Negl Trop Dis. 2012;6(5):e1659. doi: 10.1371/journal.pntd.0001659. Epub 2012 May 22. PLoS Negl Trop Dis. 2012. PMID: 22629479 Free PMC article.
-
Herpes simplex virus 1 tegument protein US11 downmodulates the RLR signaling pathway via direct interaction with RIG-I and MDA-5.J Virol. 2012 Apr;86(7):3528-40. doi: 10.1128/JVI.06713-11. Epub 2012 Feb 1. J Virol. 2012. PMID: 22301138 Free PMC article.
-
Inhibition of the type I interferon antiviral response during arenavirus infection.Viruses. 2010 Nov;2(11):2443-80. doi: 10.3390/v2112443. Epub 2010 Nov 5. Viruses. 2010. PMID: 21994626 Free PMC article.
-
Arenaviruses: Old viruses present new solutions for cancer therapy.Front Immunol. 2023 Mar 24;14:1110522. doi: 10.3389/fimmu.2023.1110522. eCollection 2023. Front Immunol. 2023. PMID: 37033933 Free PMC article. Review.
-
Function and regulation of retinoic acid-inducible gene-I.Crit Rev Immunol. 2010;30(6):489-513. doi: 10.1615/critrevimmunol.v30.i6.10. Crit Rev Immunol. 2010. PMID: 21175414 Free PMC article. Review.
References
-
- Cui, S., K. Eisenacher, A. Kirchhofer, K. Brzozka, A. Lammens, K. Lammens, T. Fujita, K. K. Conzelmann, A. Krug, and K. P. Hopfner. 2008. The C-terminal regulatory domain is the RNA 5′-triphosphate sensor of RIG-I. Mol. Cell 29:169-179. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
