

An N-terminal region of Lassa virus L protein plays a critical role in transcription but not replication of the virus genome
- PMID: 20007273
- PMCID: PMC2812395
- DOI: 10.1128/JVI.01657-09
An N-terminal region of Lassa virus L protein plays a critical role in transcription but not replication of the virus genome
Abstract
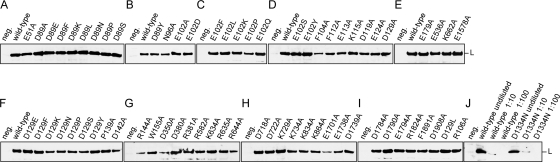
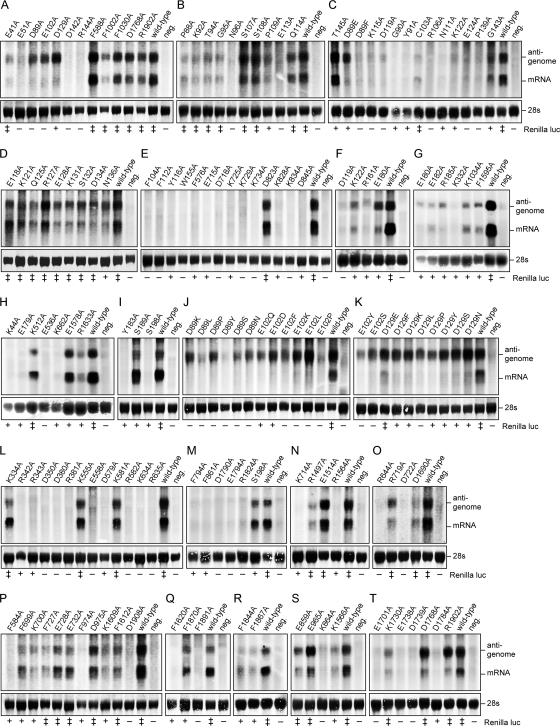
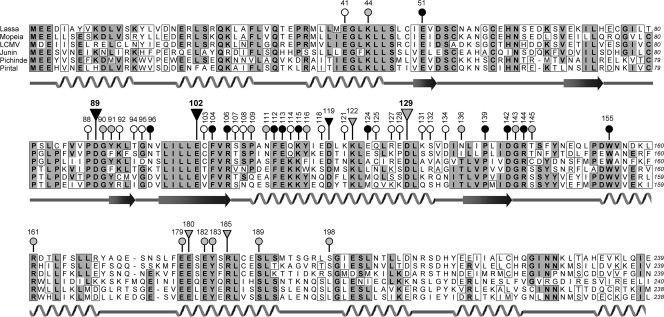
The central domain of the 200-kDa Lassa virus L protein is a putative RNA-dependent RNA polymerase. N- and C-terminal domains may harbor enzymatic functions important for viral mRNA synthesis, including capping enzymes or cap-snatching endoribonucleases. In the present study, we have employed a large-scale mutagenesis approach to map functionally relevant residues in these regions. The main targets were acidic (Asp and Glu) and basic residues (Lys and Arg) known to form catalytic and binding sites of capping enzymes and endoribonucleases. A total of 149 different mutants were generated and tested in the Lassa virus replicon system. Nearly 25% of evolutionarily highly conserved acidic and basic side chains were dispensable for function of L protein in the replicon context. The vast majority of the remaining mutants had defects in both transcription and replication. Seven residues (Asp-89, Glu-102, Asp-119, Lys-122, Asp-129, Glu-180, and Arg-185) were selectively important for mRNA synthesis. The phenotype was particularly pronounced for Asp-89, Glu-102, and Asp-129, which were indispensable for transcription but could be replaced by a variety of amino acid residues without affecting genome replication. Bioinformatics disclosed the remote similarity of this region to type IIs endonucleases. The mutagenesis was complemented by experiments with the RNA polymerase II inhibitor alpha-amanitin, demonstrating dependence of viral transcription from the cellular mRNA pool. In conclusion, this paper describes an N-terminal region in L protein being important for mRNA, but not genome synthesis. Bioinformatics and cell biological experiments lend support to the hypothesis that this region could be part of a cap-snatching enzyme.
Figures



References
-
- Buchholz, U. J., S. Finke, and K. K. Conzelmann. 1999. Generation of bovine respiratory syncytial virus (BRSV) from cDNA: BRSV NS2 is not essential for virus replication in tissue culture, and the human RSV leader region acts as a functional BRSV genome promoter. J. Virol. 73:251-259. - PMC - PubMed
-
- Dias, A., D. Bouvier, T. Crepin, A. A. McCarthy, D. J. Hart, F. Baudin, S. Cusack, and R. W. Ruigrok. 2009. The cap-snatching endonuclease of influenza virus polymerase resides in the PA subunit. Nature 458:914-918. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
