

Jasmonate-dependent and -independent pathways mediate specific effects of solar ultraviolet B radiation on leaf phenolics and antiherbivore defense
- PMID: 20007446
- PMCID: PMC2815854
- DOI: 10.1104/pp.109.148999
Jasmonate-dependent and -independent pathways mediate specific effects of solar ultraviolet B radiation on leaf phenolics and antiherbivore defense
Abstract
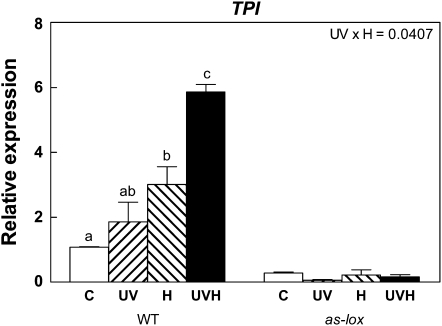
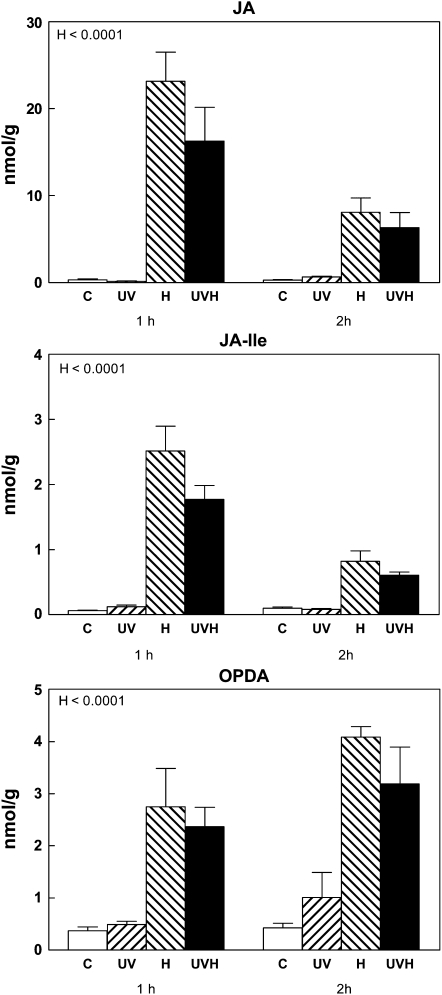
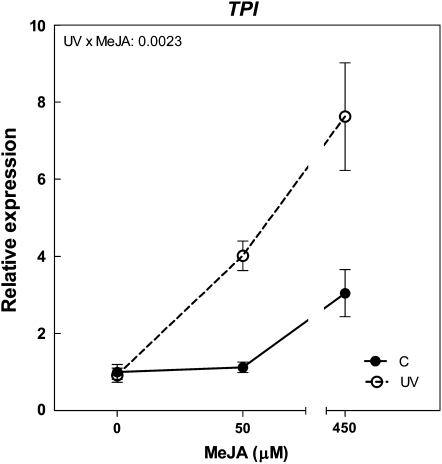
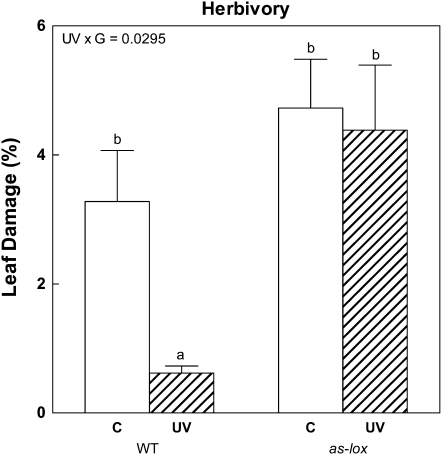
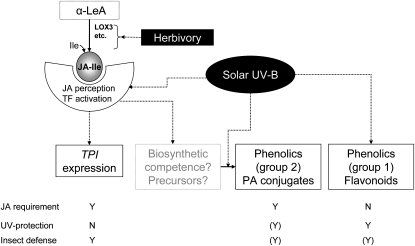
Ultraviolet B (UV-B) radiation, a very small fraction of the daylight spectrum, elicits changes in plant secondary metabolism that have large effects on plant-insect interactions. The signal transduction pathways that mediate these specific effects of solar UV-B are not known. We examined the role of jasmonate signaling by measuring responses to UV-B in wild-type and transgenic jasmonate-deficient Nicotiana attenuata plants in which a lipoxygenase gene (NaLOX3) was silenced (as-lox). In wild-type plants, UV-B failed to elicit the accumulation of jasmonic acid (JA) or the bioactive JA-isoleucine conjugate but amplified the response of jasmonate-inducible genes, such as trypsin proteinase inhibitor (TPI), to wounding and methyl jasmonate, and increased the accumulation of several phenylpropanoid derivatives. Some of these phenolic responses (accumulation of caffeoyl-polyamine conjugates) were completely lacking in as-lox plants, whereas others (accumulation of rutin and chlorogenic acid) were similar in both genotypes. In open field conditions, as-lox plants received more insect damage than wild-type plants, as expected, but the dramatic increase in resistance to herbivory elicited by UV-B exposure, which was highly significant in wild-type plants, did not occur in as-lox plants. We conclude that solar UV-B (1) uses jasmonate-dependent and -independent pathways in the elicitation of phenolic compounds, and (2) increases sensitivity to jasmonates, leading to enhanced expression of wound-response genes (TPI). The lack of UV-B-induced antiherbivore protection in as-lox plants suggests that jasmonate signaling plays a central role in the mechanisms by which solar UV-B increases resistance to insect herbivores in the field.
Figures

Similar articles
-
Jasmonates and its mimics differentially elicit systemic defence responses in Nicotiana attenuata.J Exp Bot. 2007;58(15-16):4071-82. doi: 10.1093/jxb/erm263. Epub 2007 Dec 7. J Exp Bot. 2007. PMID: 18065767
-
RuBPCase activase (RCA) mediates growth-defense trade-offs: silencing RCA redirects jasmonic acid (JA) flux from JA-isoleucine to methyl jasmonate (MeJA) to attenuate induced defense responses in Nicotiana attenuata.New Phytol. 2014 Mar;201(4):1385-1395. doi: 10.1111/nph.12591. Epub 2013 Nov 11. New Phytol. 2014. PMID: 24491116 Free PMC article.
-
Ectopic expression of AtJMT in Nicotiana attenuata: creating a metabolic sink has tissue-specific consequences for the jasmonate metabolic network and silences downstream gene expression.Plant Physiol. 2011 Sep;157(1):341-54. doi: 10.1104/pp.111.178582. Epub 2011 Jul 13. Plant Physiol. 2011. PMID: 21753114 Free PMC article.
-
The essential role of jasmonic acid in plant-herbivore interactions--using the wild tobacco Nicotiana attenuata as a model.J Genet Genomics. 2013 Dec 20;40(12):597-606. doi: 10.1016/j.jgg.2013.10.001. Epub 2013 Nov 9. J Genet Genomics. 2013. PMID: 24377866 Review.
-
Revealing insect herbivory-induced phenolamide metabolism: from single genes to metabolic network plasticity analysis.Plant J. 2014 Aug;79(4):679-92. doi: 10.1111/tpj.12503. Epub 2014 May 21. Plant J. 2014. PMID: 24617849 Free PMC article. Review.
Cited by
-
Maize response to endophytic Metarhizium robertsii is altered by water stress.PLoS One. 2023 Nov 27;18(11):e0289143. doi: 10.1371/journal.pone.0289143. eCollection 2023. PLoS One. 2023. PMID: 38011108 Free PMC article.
-
Multiple roles for UV RESISTANCE LOCUS8 in regulating gene expression and metabolite accumulation in Arabidopsis under solar ultraviolet radiation.Plant Physiol. 2013 Feb;161(2):744-59. doi: 10.1104/pp.112.211375. Epub 2012 Dec 18. Plant Physiol. 2013. PMID: 23250626 Free PMC article.
-
Protection of Photosynthesis by UVR8 and Cryptochromes in Arabidopsis Under Blue and UV Radiation.Plant Cell Environ. 2025 Aug;48(8):6321-6335. doi: 10.1111/pce.15608. Epub 2025 May 11. Plant Cell Environ. 2025. PMID: 40350778 Free PMC article.
-
The Combined Effects of Ethylene and MeJA on Metabolic Profiling of Phenolic Compounds in Catharanthus roseus Revealed by Metabolomics Analysis.Front Physiol. 2016 Jun 7;7:217. doi: 10.3389/fphys.2016.00217. eCollection 2016. Front Physiol. 2016. PMID: 27375495 Free PMC article.
-
Early changes in gene expression induced by acute UV exposure in leaves of Psychotria brachyceras, a bioactive alkaloid accumulating plant.Mol Biotechnol. 2013 May;54(1):79-91. doi: 10.1007/s12033-012-9546-3. Mol Biotechnol. 2013. PMID: 22562190
References
-
- A-H-Mackerness S, Surplus SL, Blake P, John CF, Buchanan-Wollaston V, Jordan BR, Thomas B (1999) Ultraviolet-B-induced stress and changes in gene expression in Arabidopsis thaliana: role of signalling pathways controlled by jasmonic acid, ethylene and reactive oxygen species. Plant Cell Environ 22 1413–1423
-
- Andrade A, Vigliocco A, Alemano S, Miersch O, Botella MA, Abdala G (2005) Endogenous jasmonates and octadecanoids in hypersensitive tomato mutants during germination and seedling development in response to abiotic stress. Seed Sci Res 15 309–318
-
- Ballaré CL (2009) Illuminated behaviour: phytochrome as a key regulator of light foraging and plant anti-herbivore defence. Plant Cell Environ 32 713–725 - PubMed
-
- Ballaré CL, Barnes PW, Flint SD (1995) Inhibition of hypocotyl elongation by ultraviolet-B radiation in de-etiolating tomato seedlings. 2. Time-course, comparison with flavonoid responses and adaptive significance. Physiol Plant 93 593–601
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
