

Distinct dose-dependent cortical neuronal migration and neurite extension defects in Lis1 and Ndel1 mutant mice
- PMID: 20007476
- PMCID: PMC2824645
- DOI: 10.1523/JNEUROSCI.4630-09.2009
Distinct dose-dependent cortical neuronal migration and neurite extension defects in Lis1 and Ndel1 mutant mice
Abstract
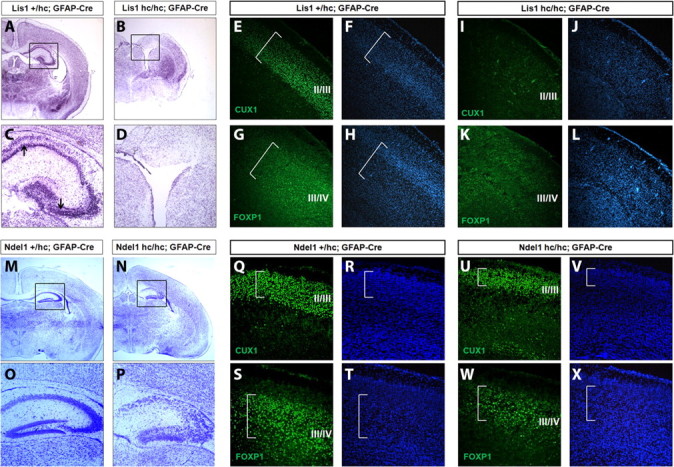
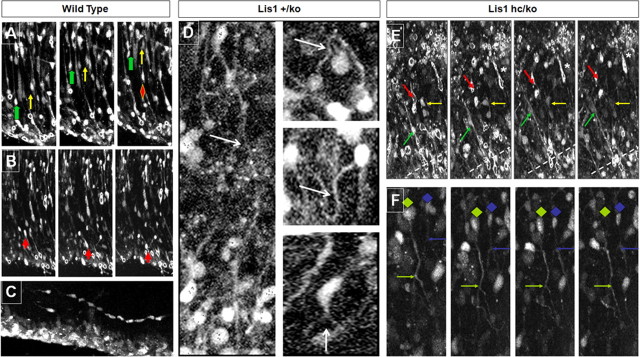
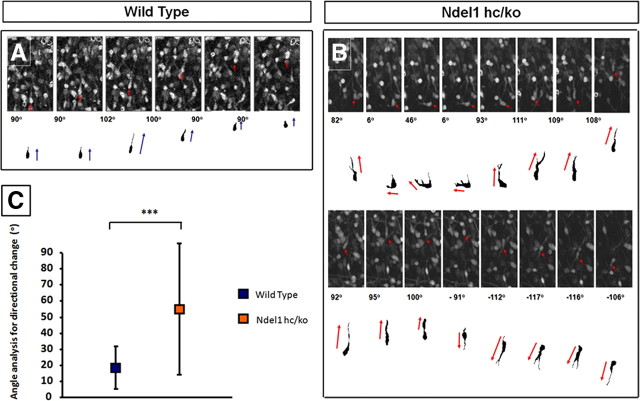
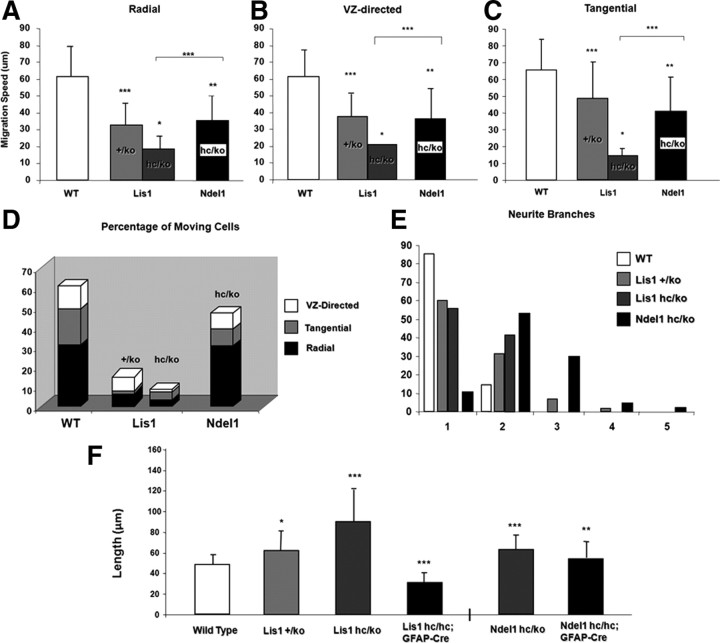
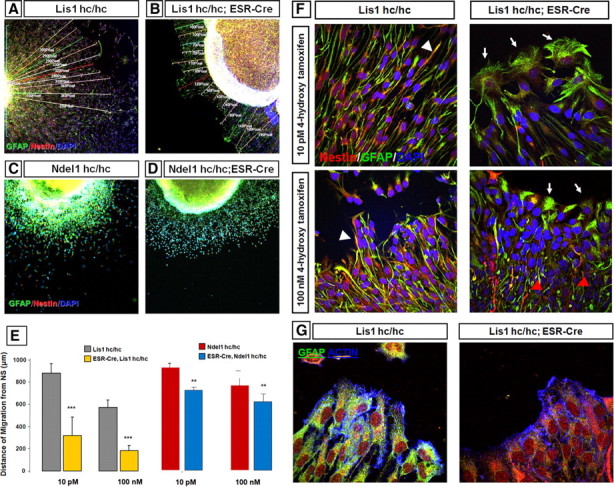
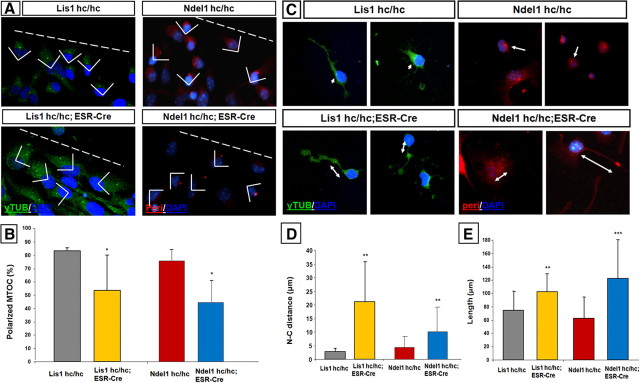
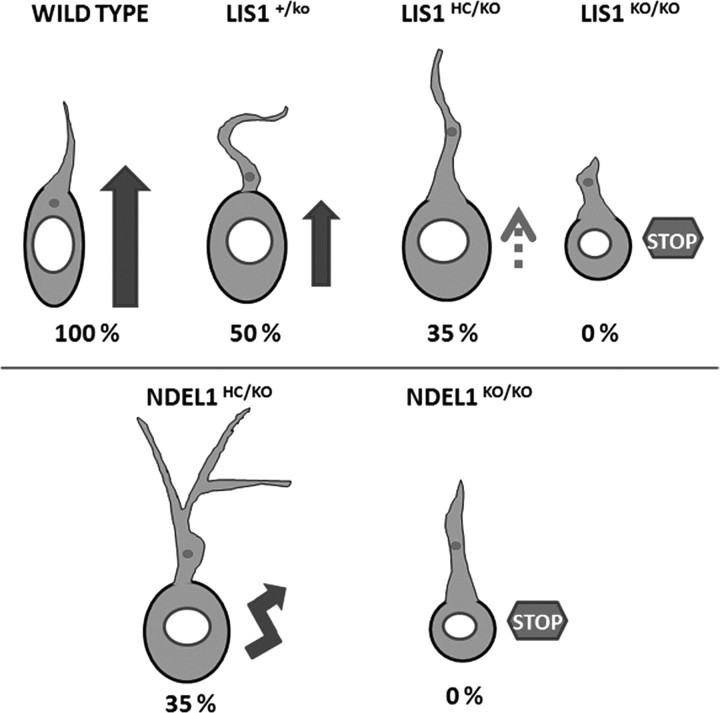
Haploinsufficiency of LIS1 results in lissencephaly, a human neuronal migration disorder. LIS1 is a microtubule- (MT) and centrosome- [microtubule organizing center (MTOC)] associated protein that regulates nucleokinesis via the regulation of dynein motor function and localization. NDEL1 (NudE isoform, NudE like) interacts with LIS1/dynein complex, and is phosphorylated by CDK5/P35. Previous reports using siRNA-mediated knock-down demonstrated similar critical roles for LIS1 and NDEL1 during neuronal migration, but neuronal migration has not been studied in genetic mutants for Lis1 and Ndel1 where protein levels are uniform in all cells. Brains from mice with complete loss of Lis1 and Ndel1 displayed severe cortical layering and hippocampal defects, but Lis1 mutants had more severe defects. Neuronal migration speed was reduced and neurite lengths were elongated in proportion to the reduction of LIS1 and NDEL1 protein levels in embryonic day 14.5 mutant cortical slices compared to wild type, using two-photon confocal time lapse videomicroscopy. Additionally, mice with 35% of wild-type NDEL1 levels displayed diverse branched migration modes with multiple leading processes, suggesting defects in adhesion and/or polarity. Complete loss of Lis1 or Ndel1 resulted in the total inhibition of nuclear movement in cortical slice assays, and in neurosphere assays, the percentage of migrating neurons with correctly polarized MTOC location was significantly reduced while nuclear-centrosomal distance was extended. Neurite lengths were increased after complete loss Ndel1 but reduced after complete loss of Lis1. Thus, Lis1 and Ndel1 are essential for normal cortical neuronal migration, neurite outgrowth, and function of the MTOC in a dose-dependent manner.
Figures

Similar articles
-
Complete loss of Ndel1 results in neuronal migration defects and early embryonic lethality.Mol Cell Biol. 2005 Sep;25(17):7812-27. doi: 10.1128/MCB.25.17.7812-7827.2005. Mol Cell Biol. 2005. PMID: 16107726 Free PMC article.
-
LIS1 controls mitosis and mitotic spindle organization via the LIS1-NDEL1-dynein complex.Hum Mol Genet. 2014 Jan 15;23(2):449-66. doi: 10.1093/hmg/ddt436. Epub 2013 Sep 12. Hum Mol Genet. 2014. PMID: 24030547 Free PMC article.
-
Novel embryonic neuronal migration and proliferation defects in Dcx mutant mice are exacerbated by Lis1 reduction.J Neurosci. 2010 Feb 24;30(8):3002-12. doi: 10.1523/JNEUROSCI.4851-09.2010. J Neurosci. 2010. PMID: 20181597 Free PMC article.
-
[Molecular mechanism of lissencephaly--how LIS1 and NDEL1 regulate cytoplasmic dynein?].Brain Nerve. 2008 Apr;60(4):375-81. Brain Nerve. 2008. PMID: 18421979 Review. Japanese.
-
[When we have learned about the brain development from a disease-oriented study: DBZ regulates cortical cell positioning and neurite extension by sustaining the anterograde transport of Lis1/DISC1 through control of Ndel1 phosphorylation].Nihon Shinkei Seishin Yakurigaku Zasshi. 2016 Apr;36(2):43-50. Nihon Shinkei Seishin Yakurigaku Zasshi. 2016. PMID: 27333658 Review. Japanese.
Cited by
-
Lis1 reduction causes tangential migratory errors in mouse spinal cord.J Comp Neurol. 2012 Apr 15;520(6):1198-211. doi: 10.1002/cne.22768. J Comp Neurol. 2012. PMID: 21935943 Free PMC article.
-
The diverse genetic landscape of neurodevelopmental disorders.Annu Rev Genomics Hum Genet. 2014;15:195-213. doi: 10.1146/annurev-genom-090413-025600. Annu Rev Genomics Hum Genet. 2014. PMID: 25184530 Free PMC article. Review.
-
Building Blocks of Functioning Brain: Cytoskeletal Dynamics in Neuronal Development.Int Rev Cell Mol Biol. 2016;322:183-245. doi: 10.1016/bs.ircmb.2015.10.002. Epub 2016 Jan 6. Int Rev Cell Mol Biol. 2016. PMID: 26940519 Free PMC article. Review.
-
Microtubule plus-end tracking proteins in neuronal development.Cell Mol Life Sci. 2016 May;73(10):2053-77. doi: 10.1007/s00018-016-2168-3. Epub 2016 Mar 11. Cell Mol Life Sci. 2016. PMID: 26969328 Free PMC article. Review.
-
Structural Consequence of Non-Synonymous Single-Nucleotide Variants in the N-Terminal Domain of LIS1.Int J Mol Sci. 2022 Mar 14;23(6):3109. doi: 10.3390/ijms23063109. Int J Mol Sci. 2022. PMID: 35328531 Free PMC article.
References
-
- Ayala R, Shu T, Tsai LH. Trekking across the brain: the journey of neuronal migration. Cell. 2007;128:29–43. - PubMed
-
- Engele J, Bohn MC. Effects of acidic and basic fibroblast growth factors (aFGF, bFGF) on glial precursor proliferation: age dependency and brain region specificity. Dev Biol. 1992;152:363–372. - PubMed
-
- Etienne-Manneville S, Hall A. Integrin-mediated activation of Cdc42 controls cell polarity in migrating astrocytes through PKCzeta. Cell. 2001;106:489–498. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous