

Neuromodulation at single presynaptic boutons of cerebellar parallel fibers is determined by bouton size and basal action potential-evoked Ca transient amplitude
- PMID: 20007482
- PMCID: PMC2829188
- DOI: 10.1523/JNEUROSCI.3793-09.2009
Neuromodulation at single presynaptic boutons of cerebellar parallel fibers is determined by bouton size and basal action potential-evoked Ca transient amplitude
Abstract
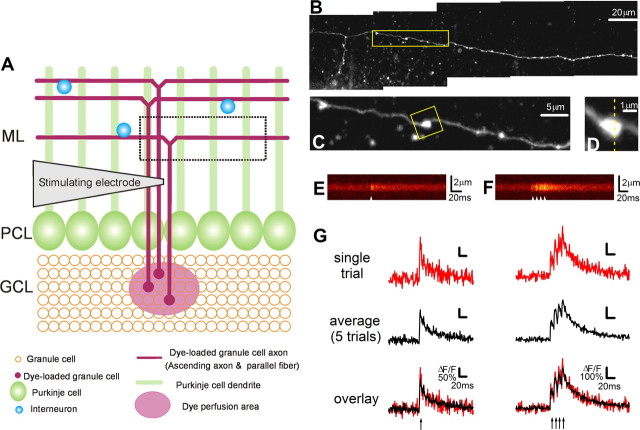
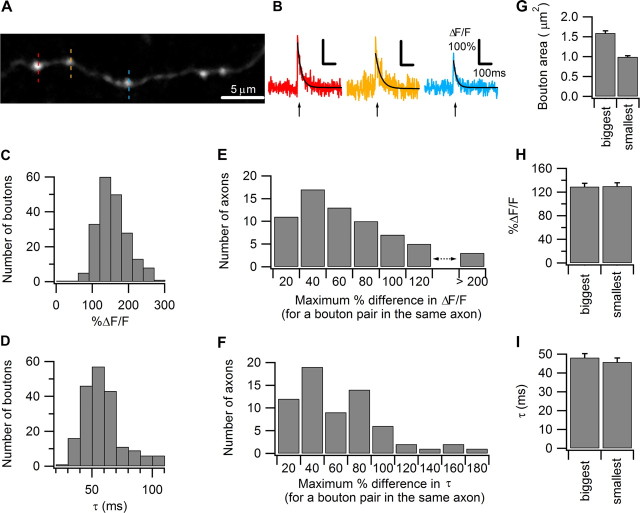
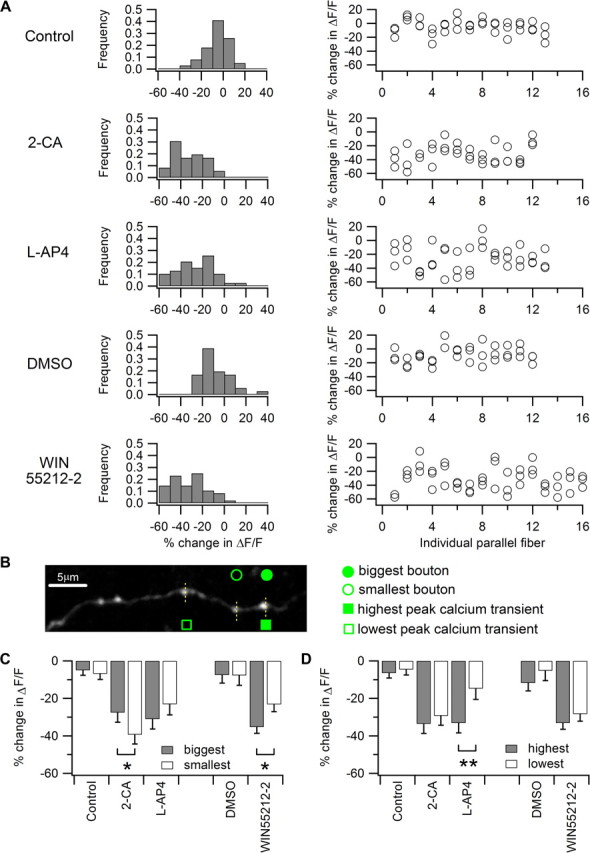
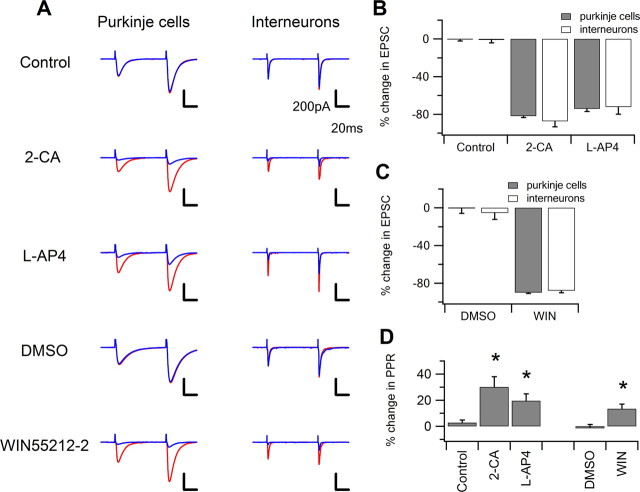
Most presynaptic terminals in the brain contain G-protein-coupled receptors that function to reduce action potential-evoked neurotransmitter release. These neuromodulatory receptors, including those for glutamate, GABA, endocannabinoids, and adenosine, exert a substantial portion of their effect by reducing evoked presynaptic Ca(2+) transients. Many axons form synapses with multiple postsynaptic neurons, but it is unclear whether presynaptic attenuation in these synapses is homogeneous, as suggested by population-level Ca(2+) imaging. We loaded Ca(2+)-sensitive dyes into cerebellar parallel fiber axons and imaged action potential-evoked Ca(2+) transients in individual presynaptic boutons with application of three different neuromodulators and found that adjacent boutons on the same axon showed striking heterogeneity in their strength of attenuation. Moreover, attenuation was predicted by bouton size or basal Ca(2+) response: smaller boutons were more sensitive to adenosine A1 agonist but less sensitive to CB1 agonist, while boutons with high basal action potential-evoked Ca(2+) transient amplitude were more sensitive to mGluR4 agonist. These results suggest that boutons within brief segment of a single parallel fiber axon can have different sensitivities toward neuromodulators and may have different capacities for both short-term and long-term plasticities.
Figures
Similar articles
-
Cannabinoid receptor modulation of synapses received by cerebellar Purkinje cells.J Neurophysiol. 2000 Mar;83(3):1167-80. doi: 10.1152/jn.2000.83.3.1167. J Neurophysiol. 2000. PMID: 10712447
-
Endocannabinoids contribute to metabotropic glutamate receptor-mediated inhibition of GABA release onto hippocampal CA3 pyramidal neurons in an isolated neuron/bouton preparation.Neuroscience. 2010 Feb 17;165(4):1377-89. doi: 10.1016/j.neuroscience.2009.11.054. Epub 2009 Dec 1. Neuroscience. 2010. PMID: 19961906
-
Properties of 4 Hz stimulation-induced parallel fiber-Purkinje cell presynaptic long-term plasticity in mouse cerebellar cortex in vivo.Eur J Neurosci. 2014 May;39(10):1624-31. doi: 10.1111/ejn.12559. Epub 2014 Mar 26. Eur J Neurosci. 2014. PMID: 24666426
-
Calcium influx measured at single presynaptic boutons of cerebellar granule cell ascending axons and parallel fibers.Cerebellum. 2012 Mar;11(1):121-31. doi: 10.1007/s12311-009-0151-3. Cerebellum. 2012. PMID: 20049574 Free PMC article.
-
Endocannabinoids inhibit transmission at granule cell to Purkinje cell synapses by modulating three types of presynaptic calcium channels.J Neurosci. 2004 Jun 16;24(24):5623-31. doi: 10.1523/JNEUROSCI.0918-04.2004. J Neurosci. 2004. PMID: 15201335 Free PMC article.
Cited by
-
A new signalling pathway for parallel fibre presynaptic type 4 metabotropic glutamate receptors (mGluR4) in the rat cerebellar cortex.J Physiol. 2012 Jul 1;590(13):2977-94. doi: 10.1113/jphysiol.2012.232074. Epub 2012 May 8. J Physiol. 2012. PMID: 22570379 Free PMC article.
-
Long-term depression of presynaptic cannabinoid receptor function at parallel fibre synapses.J Physiol. 2019 Jun;597(12):3167-3181. doi: 10.1113/JP277727. Epub 2019 May 13. J Physiol. 2019. PMID: 31020998 Free PMC article.
-
Presynaptic calcium channel inhibition underlies CB₁ cannabinoid receptor-mediated suppression of GABA release.J Neurosci. 2014 Jun 4;34(23):7958-63. doi: 10.1523/JNEUROSCI.0247-14.2014. J Neurosci. 2014. PMID: 24899717 Free PMC article.
-
An evolving view of retinogeniculate transmission.Vis Neurosci. 2017 Jan;34:E013. doi: 10.1017/S0952523817000104. Vis Neurosci. 2017. PMID: 28965513 Free PMC article. Review.
-
Release properties of individual presynaptic boutons expressed during homosynaptic depression and heterosynaptic facilitation of the Aplysia sensorimotor synapse.Front Cell Neurosci. 2013 Sep 24;7:165. doi: 10.3389/fncel.2013.00165. eCollection 2013. Front Cell Neurosci. 2013. PMID: 24068986 Free PMC article.
References
-
- Abitbol K, Acher F, Daniel H. Depression of excitatory transmission at PF-PC synapse by group III metabotropic glutamate receptors is provided exclusively by mGluR4 in the rodent cerebellar cortex. J Neurochem. 2008;105:2069–2079. - PubMed
-
- Bailey CH, Chen M. Morphological basis of long-term habituation and sensitization in Aplysia. Science. 1983;220:91–93. - PubMed
-
- Batchelor AM, Garthwaite J. GABAB receptors in the parallel fibre pathway of rat cerebellum. Eur J Neurosci. 1992;4:1059–1064. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous