

GTPase-Rac enhances depolarization-induced superoxide production by the macula densa during tubuloglomerular feedback
- PMID: 20007513
- PMCID: PMC2828178
- DOI: 10.1152/ajpregu.00622.2009
GTPase-Rac enhances depolarization-induced superoxide production by the macula densa during tubuloglomerular feedback
Abstract
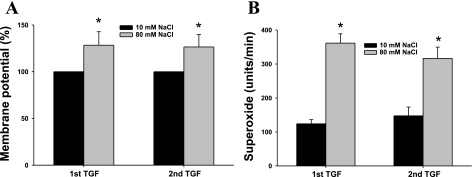
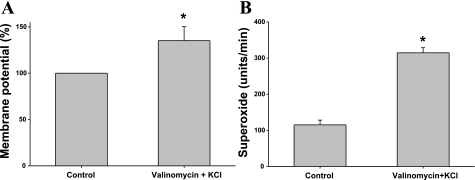
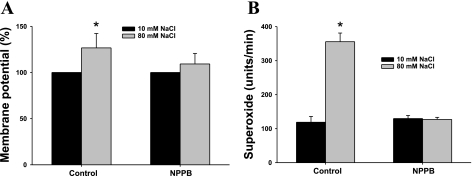
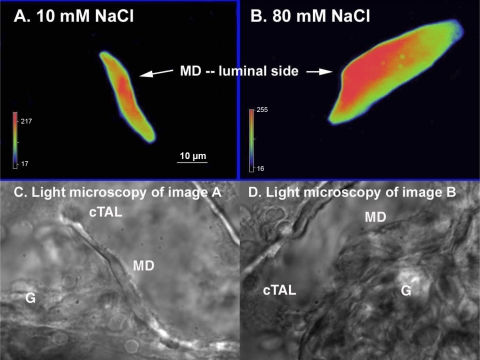
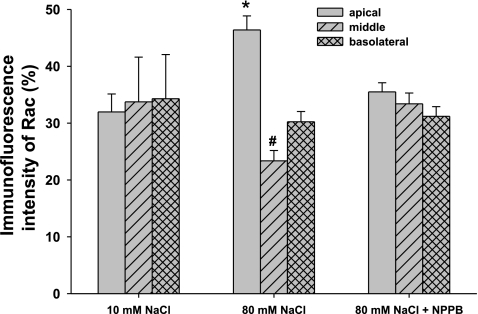
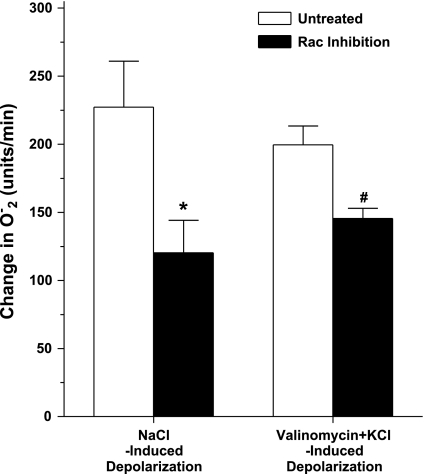
Superoxide (O(2)(-) ) enhances tubuloglomerular feedback (TGF) by scavenging nitric oxide at the macula densa (MD). The primary source of O(2)(-) in the MD during TGF is NADPH oxidase, which is activated by membrane depolarization. While Rac, a small GTP-binding protein, has been shown to enhance NADPH oxidase activity, its role in O(2)(-) generation by the MD is unknown. We hypothesized that depolarization of the MD leads to translocation of Rac to the apical membrane, and its activation, in turn, augments O(2)(-) generation during TGF. We tested this by measuring membrane potential and increased O(2)(-) levels during TGF responses in isolated, perfused tubules containing the intact MD plaque. Switching tubular NaCl from 10 to 80 mM, which induces TGF, depolarized membrane potential by 28.4 + or - 4.5% from control (P < 0.05) and O(2)(-) levels from 124 + or - 19 to 361 + or - 27 U/min. This NaCl-induced depolarization and O(2)(-) generation were blocked by a Cl(-) channel blocker, 5-nitro-2(3-phenylpropylamino) benzoic acid (NPPB; 10(-6) M). Inhibition of Rac blunted NaCl-induced O(2)(-) generation by 47%. When the NaCl content of the MD perfusate was increased from 10 to 80 mM, immunointensity of Rac on the apical side increased from 32 + or - 3.1 to 46 + or - 2.5% of the total immunofluorescence in the MD, indicating that high NaCl induces the translocation of Rac to the apical membrane. This NaCl-induced Rac translocation was blocked by a Cl(-) channel blocker, NPPB, indicating that depolarization of the MD induced Rac translocation. In conclusion, we found that depolarization of the MD during TGF leads to translocation of Rac to the apical membrane, which enhances O(2)(-) generation by the MD.
Figures
References
-
- Bell PD, Lapointe JY, Peti-Peterdi J. Macula densa cell signaling. Annu Rev Physiol 65: 32.1–32.20, 2003 - PubMed
-
- Bell PD, Lapointe JY, Cardinal J. Direct measurement of basolateral membrane potentials from cells of the macula densa. Am J Physiol Renal Fluid Electrolyte Physiol 257: F463–F468, 1989 - PubMed
-
- Briggs JP, Schnermann J. Macula densa control of renin secretion and glomerular vascular tone: evidence for common cellular mechanisms. Renal Physiol (Basel) 9: 193–203, 1986 - PubMed
-
- Cheng G, Diebold BA, Hughes Y, Lambeth JD. Nox1-dependent reactive oxygen generation is regulated by Rac1. J Biol Chem 281: 17718–17726, 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous