

Review
doi: 10.1056/NEJMra0903373.
Telomere diseases
Affiliations
- PMID: 20007561
- PMCID: PMC3401586
- DOI: 10.1056/NEJMra0903373
Item in Clipboard
Review
Telomere diseases
N Engl J Med.
.
No abstract available
Conflict of interest statement
No potential conflict of interest relevant to this article was reported.
Figures
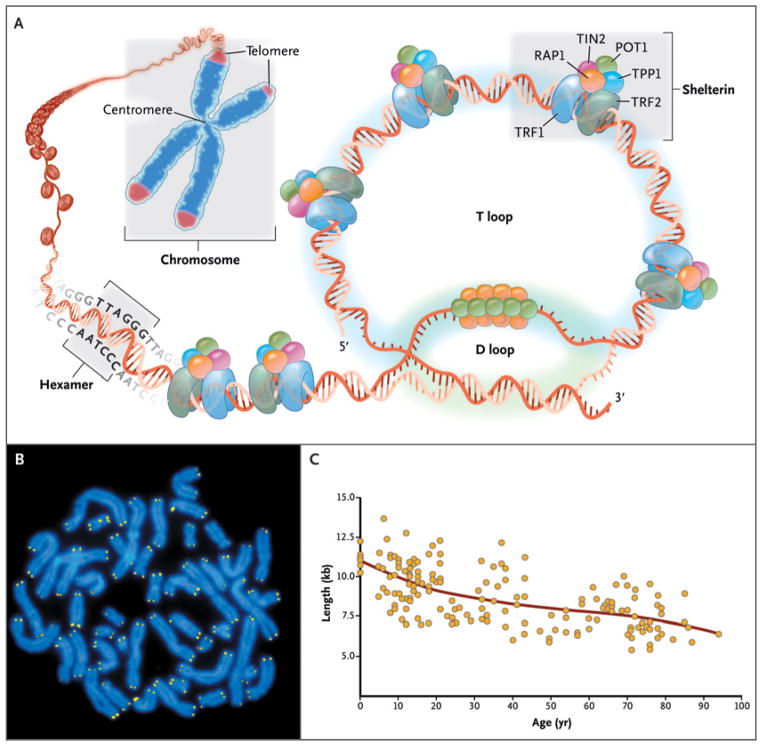
As shown in Panel A, telomeres are located at the ends of linear chromosomes; they are composed of hundreds to thousands of tandem DNA repeat sequences: hexameric TTAGGG in the leading strand and CCCTAA in the lagging strand in humans. Protective proteins associated with telomere DNA are collectively termed shelterin (TRF1, TRF2, TIN2, POT1, TPP1, and RAP1). The 3′ end of the telomeric leading strand terminates as a single-stranded overhang, which folds back and invades the double-stranded telomeric helix, forming the T loop. As shown in Panel B, telomeres can be directly visualized under the microscope at the ends of metaphase chromosomes (four telomere signals per chromosome) by fluorescence in situ hybridization (FISH). (Image provided by Peter Lansdorp, M.D., Ph.D.) Average telomere length can be measured by several methods: a technique that combines flow cytometry and FISH (flow-FISH), Southern blotting, and a quantitative polymerase-chain-reaction (qPCR) assay. Flow-FISH can measure the telomere length in different cell subgroups, such as granulocytes or CD4+ T lymphocytes; Southern blotting reveals length and length heterogeneity; and qPCR is a rapid assay that requires very small amounts of DNA. As shown in Panel C, the average length of telomeres in human leukocytes varies, ranging from approximately 11 kb at birth (in umbilical-cord blood) to 6 kb at 90 years of age. Telomere loss is most rapid early in life, and over a life span it is not linear but follows a third-order polynomial. Data are from Yamaguchi et al.
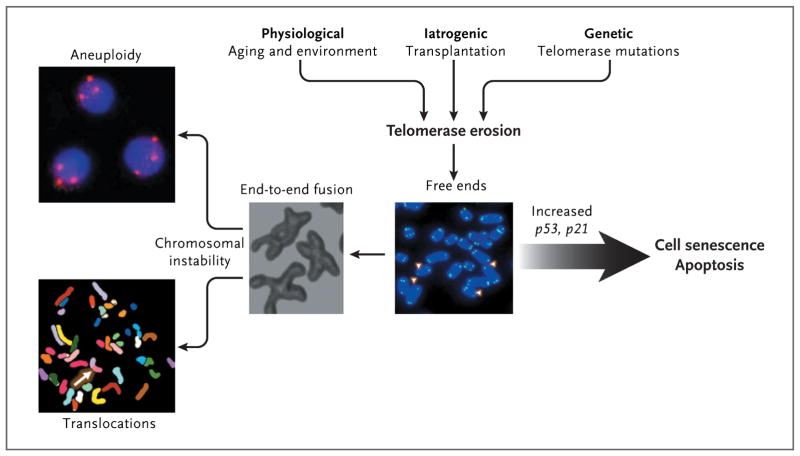
Telomeres inexorably shorten with every cell division, and telomere attrition is an inevitable physiological consequence of aging. Telomere shortening also may be iatrogenic; for example, telomere shortening occurs after bone marrow transplantation, in which highly proliferative hematopoietic stem cells and progenitor cells reconstitute hematopoiesis. Environmental factors also may accelerate telomere loss. In addition, telomere attrition may be genetic; there may be an inherited inability to elongate telomeres as a result of mutations in components of the telomerase complex. When telomeres become critically short, inappropriately capped chromosomes or telomere-free ends emerge, which lead to cell senescence or apoptosis. If the cell overrides senescence and continues to proliferate (e.g., because of inactive p53), uncapped telomeres may cause end-to-end fusion of chromosomes, breakage-fusion-bridge cycles, aneuploidy, and chromosomal translocations.
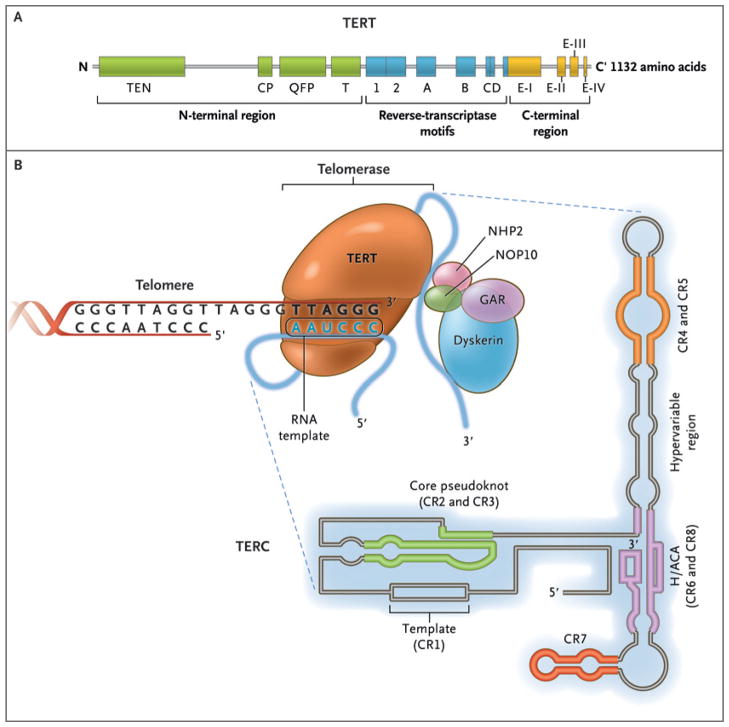
The enzyme telomerase reverse transcriptase (TERT), its RNA component (TERC), the protein dyskerin, and other associated proteins (NHP2, NOP10, and GAR1) are shown. Telomerase catalytically adds TTAGGG hexameric nucleotide repeats to the 3′-hydroxyl end of the telomeric leading strand, using a specific sequence in the RNA component as the template. TERT contains three major domains: the N-terminal region, the reverse-transcriptase motifs, and the C-terminal region, all containing evolutionarily conserved motifs. TERC contains 451 nucleotides in seven conserved regions (CR1 through CR7), including the template (CR1), and an H/ACA box, a hairpin nucleotide sequence characteristic of a class of small nucleolar RNAs involved in RNA processing.
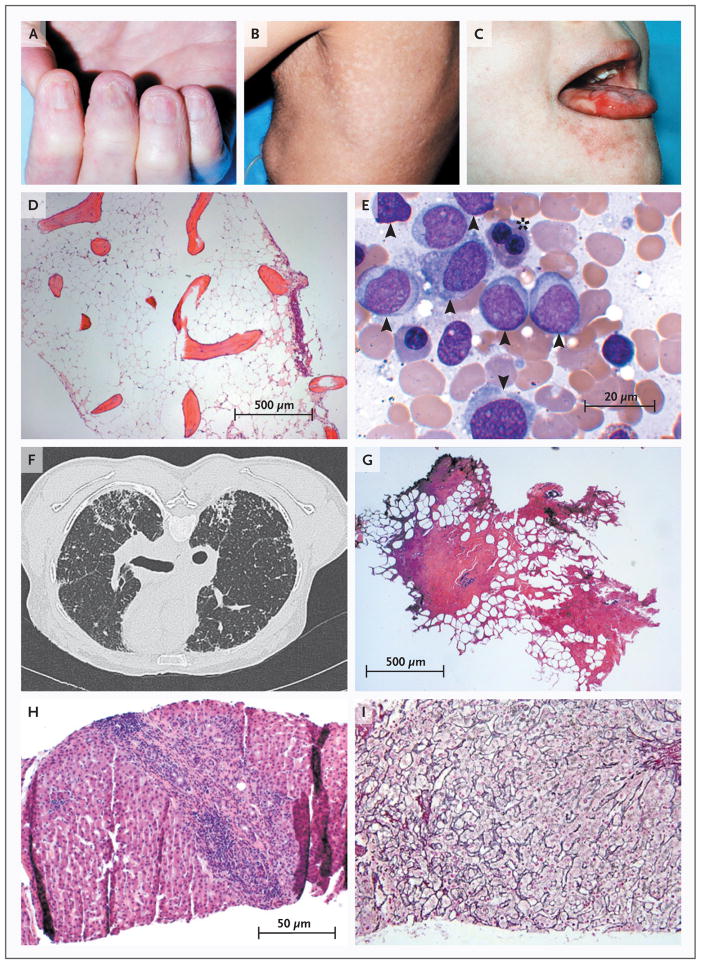
The childhood syndrome dyskeratosis congenita is characterized clinically by the mucocutaneous triad of nail dystrophy (Panel A), reticular skin hyperpigmentation or hypopigmentation (Panel B), and leukoplakia (Panel C) (reprinted from Alter, with the permission of the publisher). In the bone marrow, telomere shortening confers a predisposition to aplastic anemia (Panel D, Giemsa stain) and progression to myelodysplasia and acute myeloid leukemia (Panel E, Giemsa stain). Leukemic bone marrow is characterized by an increased number of myeloid blasts (arrowheads) and dysplasia or dyserythropoiesis (asterisk). In the lungs, telomere attrition can be clinically manifested as pulmonary fibrosis and radiologically characterized by diffuse fibrosis predominantly in the subpleural region (Panel F). Histologically, fibrotic zones alternate with less affected parenchyma (Panel G, hematoxylin and eosin). Telomere shortening in the liver has diverse histologic appearances, including cirrhosis with inflammation (Panel H, hematoxylin and eosin) and nodular regenerative hyperplasia (Panel I, reticulin stain).
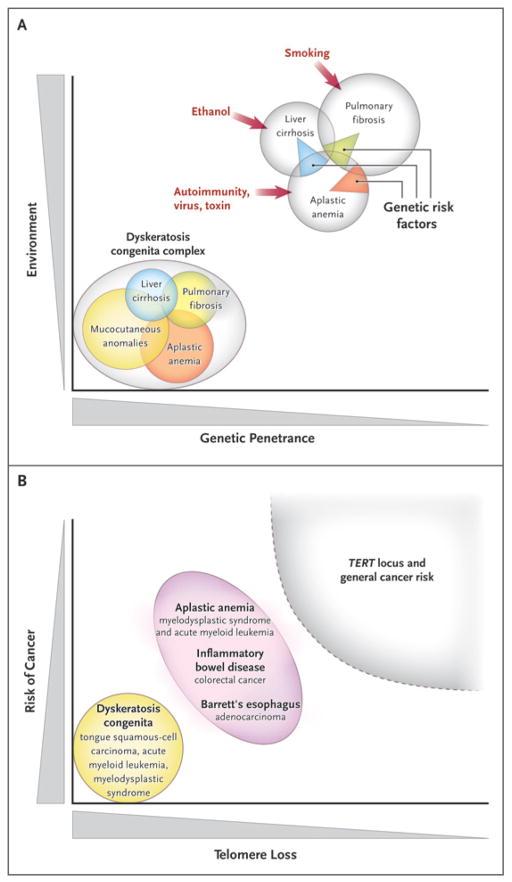
Panel A shows a Venn diagram of mutations of the telomerase complex and human telomere diseases. Dyskeratosis congenita is the most evident and severe manifestation of genetic lesions causing telomere diseases, with high genetic penetrance and congenital clinical manifestations. However, telomerase mutations may be less penetrant and induce single-organ damage in adults without suggestive family histories and the classic physical signs of dyskeratosis congenita. Thus, telomerase mutations represent risk factors rather than genetic determinants in aplastic anemia, pulmonary fibrosis, and liver cirrhosis. Environmental, epigenetic, and other genetic factors probably contribute to disease development in these patients. Panel B shows the relationship between telomere shortening and the risk of cancer. In dyskeratosis congenita, in which genetic penetrance is high, the risk of the development of cancer — particularly head and neck squamous-cell carcinoma and acute myeloid leukemia — also is elevated. In addition, patients with aplastic anemia are at risk for the development of clonal malignant disorders, but the risk is lower than that among patients with dyskeratosis congenita. Similarly, short telomeres appear to predict the progression of chronic inflammatory gastrointestinal states to adenocarcinoma. In multiple genomewide association studies, the TERT locus has appeared as a significant susceptibility locus for a variety of cancers, but at relatively low odds ratios. Shaded areas representing diseases and disease states are not drawn to scale.
Comment in
-
Telomere diseases.N Engl J Med. 2010 Mar 25;362(12):1150. doi: 10.1056/NEJMc1000801. N Engl J Med. 2010. PMID: 20335598 No abstract available.
References
-
- Szostak JW, Blackburn EH. Cloning yeast telomeres on linear plasmid vectors. Cell. 1982;29:245–55. - PubMed
-
- Greider CW, Blackburn EH. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell. 1985;43:405–13. - PubMed
-
- Heiss NS, Knight SW, Vulliamy TJ, et al. X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions. Nat Genet. 1998;19:32–8. - PubMed
-
- Mitchell JR, Wood E, Collins K. A telo-merase component is defective in the human disease dyskeratosis congenita. Nature. 1999;402:551–5. - PubMed
-
- Olovnikov AM. Principle of marginotomy in template synthesis of polynucleotides. Dokl Akad Nauk SSSR. 1971;201:1496–9. (In Russian.) - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical