

Membrane association of mitochondrial DNA facilitates base excision repair in mammalian mitochondria
- PMID: 20007607
- PMCID: PMC2836570
- DOI: 10.1093/nar/gkp1143
Membrane association of mitochondrial DNA facilitates base excision repair in mammalian mitochondria
Abstract
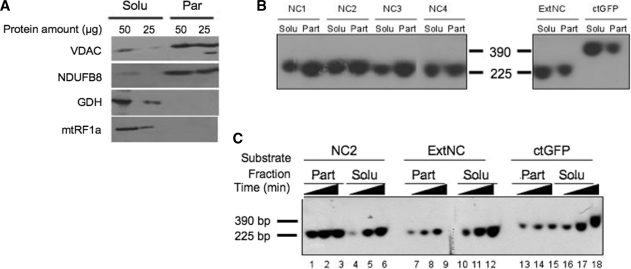
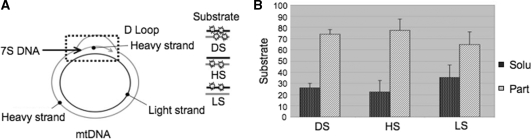
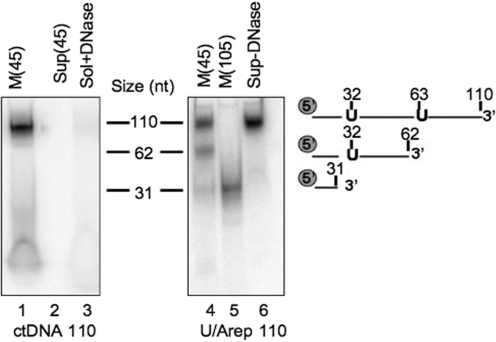
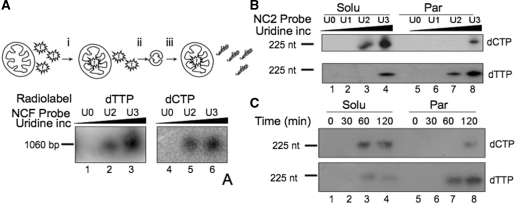
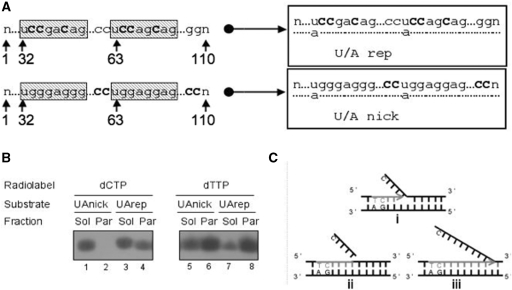
Mitochondrial DNA encodes a set of 13 polypeptides and is subjected to constant oxidative stress due to ROS production within the organelle. It has been shown that DNA repair in the mitochondrion proceeds through both short- and long-patch base excision repair (BER). In the present article, we have used the natural competence of mammalian mitochondria to import DNA and study the sub-mitochondrial localization of the repair system in organello. Results demonstrate that sequences corresponding to the mtDNA non-coding region interact with the inner membrane in a rapid and saturable fashion. We show that uracil containing import substrates are taken into the mitochondrion and are used as templates for damage driven DNA synthesis. After further sub-fractionation, we show that the length of the repair synthesis patch differs in the soluble and the particulate fraction. Bona fide long patch BER synthesis occurs on the DNA associated with the particulate fraction, whereas a nick driven DNA synthesis occurs when the uracil containing DNA accesses the soluble fraction. Our results suggest that coordinate interactions of the different partners needed for BER is only found at sites where the DNA is associated with the membrane.
Figures
References
-
- Hudson EK, Hogue BA, Souza-Pinto NC, Croteau DL, Anson RM, Bohr VA, Hansford RG. Age-associated change in mitochondrial DNA damage. Free Radic. Res. 1998;29:573–579. - PubMed
-
- Trifunovic A, Wredenberg A, Falkenberg M, Spelbrink JN, Rovio AT, Bruder CE, Bohlooly YM, Gidlof S, Oldfors A, Wibom R, et al. Premature ageing in mice expressing defective mitochondrial DNA polymerase. Nature. 2004;429:417–423. - PubMed
