

A mutation in GDP-mannose pyrophosphorylase causes conditional hypersensitivity to ammonium, resulting in Arabidopsis root growth inhibition, altered ammonium metabolism, and hormone homeostasis
- PMID: 20007685
- PMCID: PMC2803207
- DOI: 10.1093/jxb/erp310
A mutation in GDP-mannose pyrophosphorylase causes conditional hypersensitivity to ammonium, resulting in Arabidopsis root growth inhibition, altered ammonium metabolism, and hormone homeostasis
Abstract
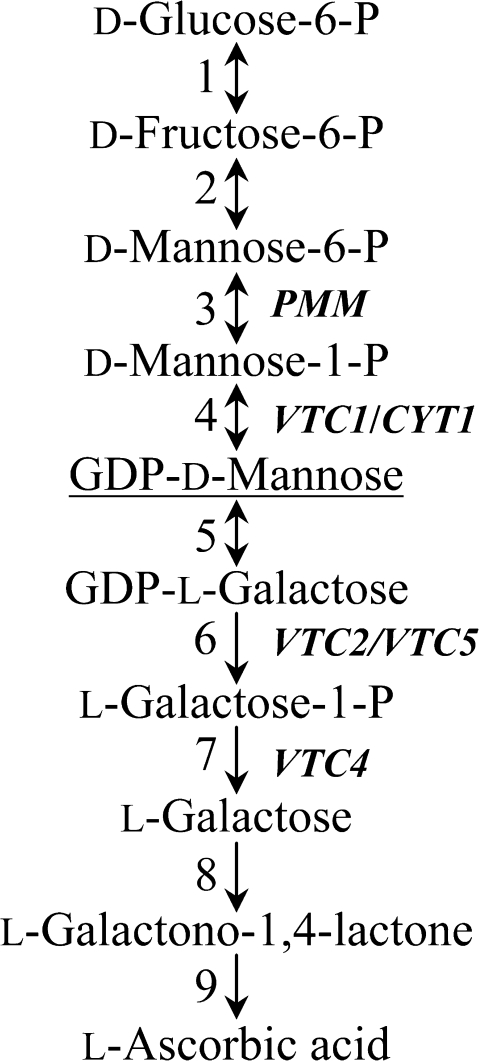
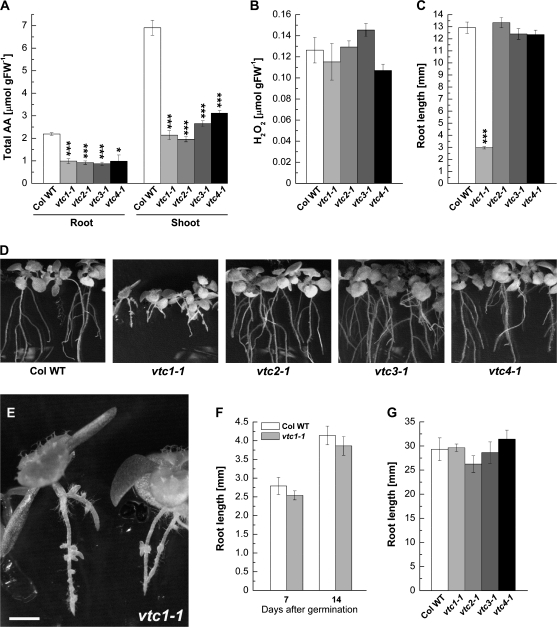
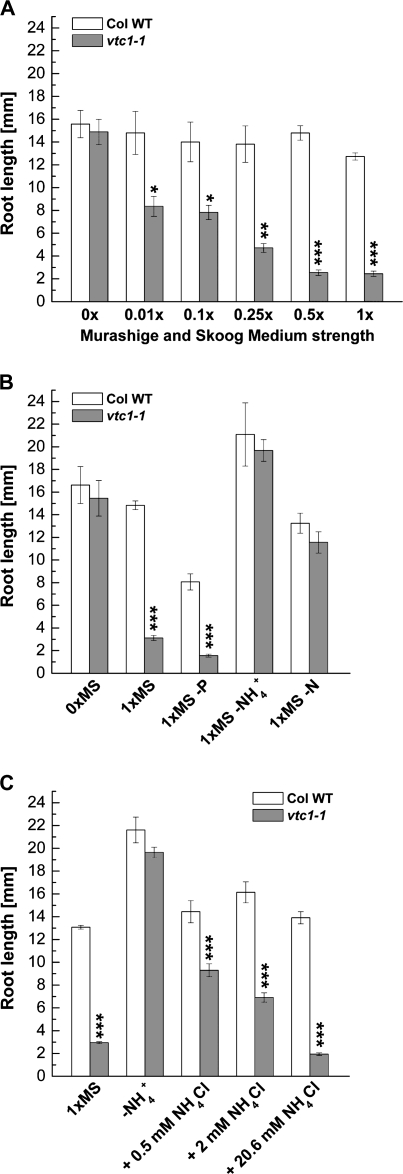
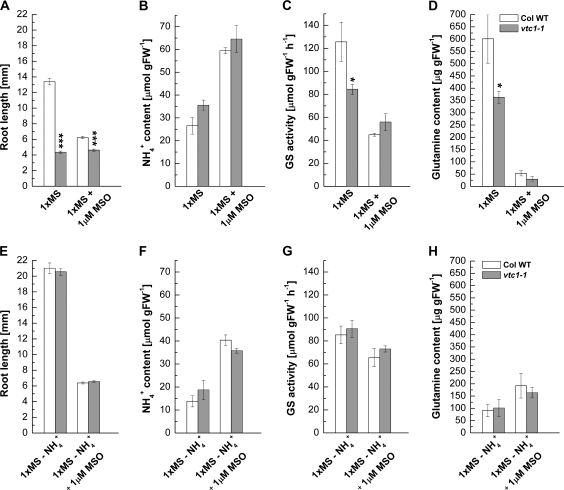
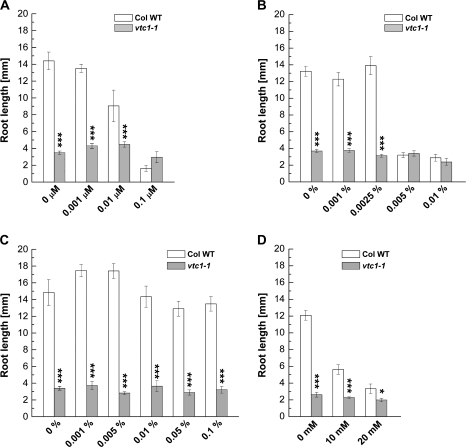
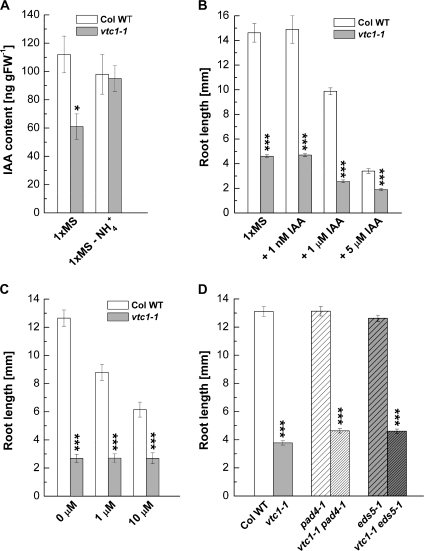
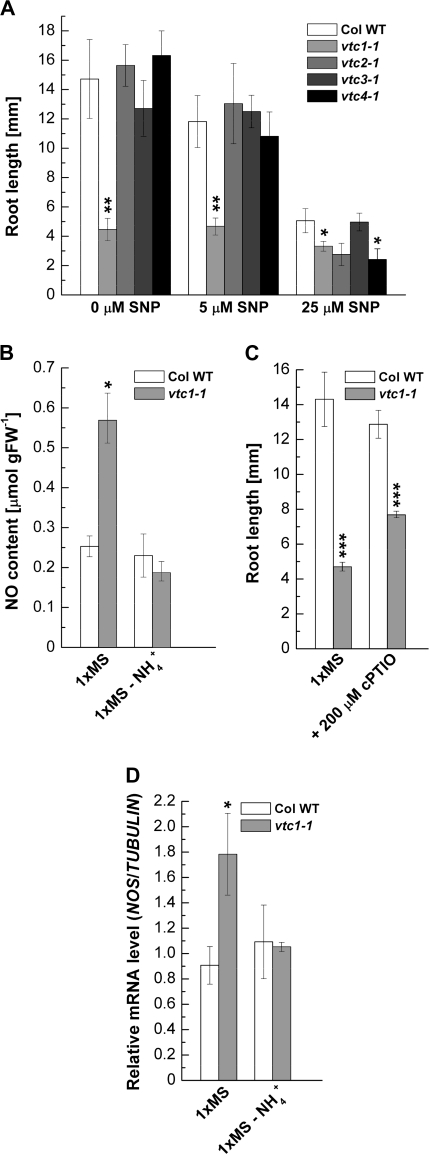
Ascorbic acid (AA) is an antioxidant fulfilling a multitude of cellular functions. Given its pivotal role in maintaining the rate of cell growth and division in the quiescent centre of the root, it was hypothesized that the AA-deficient Arabidopsis thaliana mutants vtc1-1, vtc2-1, vtc3-1, and vtc4-1 have altered root growth. To test this hypothesis, root development was studied in the wild type and vtc mutants grown on Murashige and Skoog medium. It was discovered, however, that only the vtc1-1 mutant has strongly retarded root growth, while the other vtc mutants exhibit a wild-type root phenotype. It is demonstrated that the short-root phenotype in vtc1-1 is independent of AA deficiency and oxidative stress. Instead, vtc1-1 is conditionally hypersensitive to ammonium (NH(4)(+)). To provide new insights into the mechanism of NH(4)(+) sensitivity in vtc1-1, root development, NH(4)(+) content, glutamine synthetase (GS) activity, glutamate dehydrogenase activity, and glutamine content were assessed in wild-type and vtc1-1 mutant plants grown in the presence and absence of high NH(4)(+) and the GS inhibitor MSO. Since VTC1 encodes a GDP-mannose pyrophosphorylase, an enzyme generating GDP-mannose for AA biosynthesis and protein N-glycosylation, it was also tested whether protein N-glycosylation is affected in vtc1-1. Furthermore, since root development requires the action of a variety of hormones, it was investigated whether hormone homeostasis is linked to NH(4)(+) sensitivity in vtc1-1. Our data suggest that NH(4)(+) hypersensitivity in vtc1-1 is caused by disturbed N-glycosylation and that it is associated with auxin and ethylene homeostasis and/or nitric oxide signalling.
Figures
References
-
- Barker AV. Ammonium accumulation and ethylene evolution by tomato infected with root-knot nematode and grown under different regimes of plant nutrition. Communications in Soil Science and Plant Analysis. 1999a;30:175–182.
-
- Barker AV. Foliar ammonium accumulation as an index of stress in plants. Communications in Soil Science and Plant Analysis. 1999b;30:167–174.
-
- Bhalerao RP, Eklof J, Ljung K, Marchant A, Bennett M, Sandberg G. Shoot-derived auxin is essential for early lateral root emergence in Arabidopsis seedlings. The Plant Journal. 2002;29:325–332. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
