

miR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis
- PMID: 20007771
- PMCID: PMC2799693
- DOI: 10.1073/pnas.0908718106
miR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis
Abstract
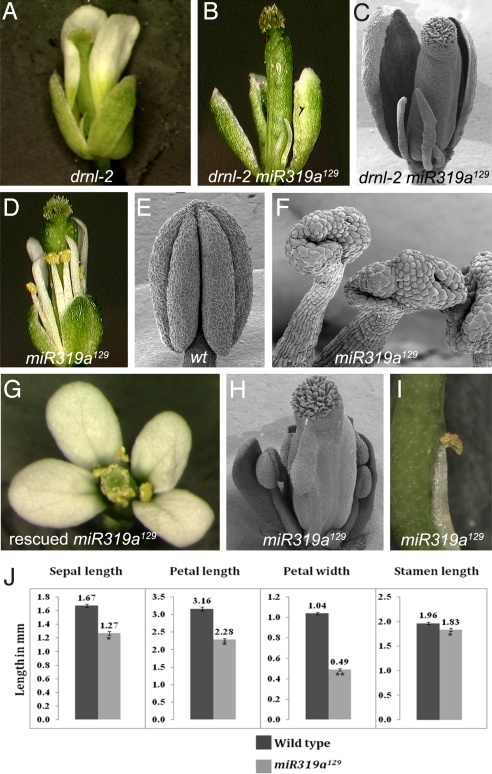
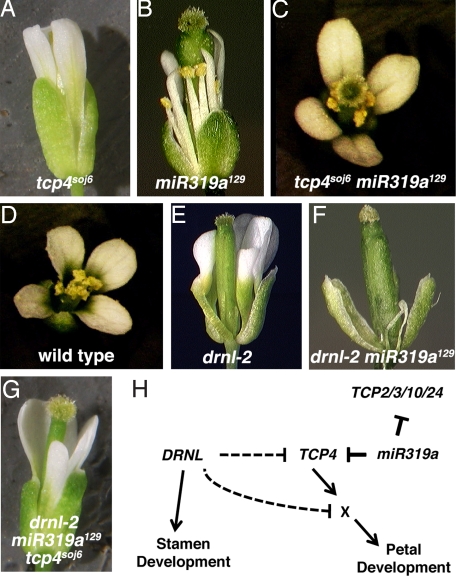
In a genetic screen in a drnl-2 background, we isolated a loss-of-function allele in miR319a (miR319a(129)). Previously, miR319a has been postulated to play a role in leaf development based on the dramatic curled-leaf phenotype of plants that ectopically express miR319a (jaw-D). miR319a(129) mutants exhibit defects in petal and stamen development; petals are narrow and short, and stamens exhibit defects in anther development. The miR319a(129) loss-of-function allele contains a single-base change in the middle of the encoded miRNA, which reduces the ability of miR319a to recognize targets. Analysis of the expression patterns of the three members of the miR319 gene family (miR319a, miR319b, and miR319c) indicates that these genes have largely non-overlapping expression patterns suggesting that these genes have distinct developmental functions. miR319a functions by regulating the TCP transcription factors TCP2, TCP3, TCP4, TCP10, and TCP24; the level of RNA expression of these TCP genes is down-regulated in jaw-D and elevated in miR319a(129). Several lines of evidence demonstrate that TCP4 is a key target of miR319a. First, the tcp4(soj6) mutant, which contains a mutation in the TCP4 miRNA-binding site complementary to the miR319a(129) mutation, suppresses the flower phenotype of miR319a(129). Second, expression of wild-type TCP4 in petals and stamens (i.e., AP3:TCP4) has no effect on flower development; by contrast, a miRNA-resistant version of TCP4, when expressed in petals and stamens (i.e., pAP3:mTCP4) causes these organs not to develop. Surprisingly, when AP3:TCP4 is present in a miR319a(129) background, petal and stamen development is severely disrupted, suggesting that proper regulation by miR319a of TCP4 is critical in these floral organs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Spatial Control of Gene Expression by miR319-Regulated TCP Transcription Factors in Leaf Development.Plant Physiol. 2018 Feb;176(2):1694-1708. doi: 10.1104/pp.17.00823. Epub 2017 Nov 13. Plant Physiol. 2018. PMID: 29133375 Free PMC article.
-
RABBIT EARS regulates the transcription of TCP4 during petal development in Arabidopsis.J Exp Bot. 2016 Dec;67(22):6473-6480. doi: 10.1093/jxb/erw419. Epub 2016 Nov 12. J Exp Bot. 2016. PMID: 27838638 Free PMC article.
-
Inhibition of Arabidopsis thaliana CIN-like TCP transcription factors by Agrobacterium T-DNA-encoded 6B proteins.Plant J. 2020 Mar;101(6):1303-1317. doi: 10.1111/tpj.14591. Epub 2019 Dec 5. Plant J. 2020. PMID: 31659801 Free PMC article.
-
Regulation of reproductive development by non-coding RNA in Arabidopsis: to flower or not to flower.J Plant Res. 2012 Nov;125(6):693-704. doi: 10.1007/s10265-012-0513-7. Epub 2012 Jul 27. J Plant Res. 2012. PMID: 22836383 Free PMC article. Review.
-
Building beauty: Understanding how hormone signaling regulates petal patterning and morphogenesis.Plant J. 2025 Mar;121(6):e70101. doi: 10.1111/tpj.70101. Plant J. 2025. PMID: 40106266 Free PMC article. Review.
Cited by
-
Constitutive expression of a miR319 gene alters plant development and enhances salt and drought tolerance in transgenic creeping bentgrass.Plant Physiol. 2013 Mar;161(3):1375-91. doi: 10.1104/pp.112.208702. Epub 2013 Jan 4. Plant Physiol. 2013. PMID: 23292790 Free PMC article.
-
Jasmonic acid protects etiolated seedlings of Arabidopsis thaliana against herbivorous arthropods.Plant Signal Behav. 2016 Aug 2;11(8):e1214349. doi: 10.1080/15592324.2016.1214349. Plant Signal Behav. 2016. PMID: 27485473 Free PMC article.
-
Physiological and transcriptional responses of two contrasting Populus clones to nitrogen stress.Tree Physiol. 2016 May;36(5):628-42. doi: 10.1093/treephys/tpw019. Epub 2016 Apr 19. Tree Physiol. 2016. PMID: 27095258 Free PMC article.
-
Functional intron-derived miRNAs and host-gene expression in plants.Plant Methods. 2018 Sep 24;14:83. doi: 10.1186/s13007-018-0351-2. eCollection 2018. Plant Methods. 2018. PMID: 30258486 Free PMC article.
-
Identification of miRNA from Porphyra yezoensis by high-throughput sequencing and bioinformatics analysis.PLoS One. 2010 May 19;5(5):e10698. doi: 10.1371/journal.pone.0010698. PLoS One. 2010. PMID: 20502668 Free PMC article.
References
-
- Garcia D. A miRacle in plant development: Role of microRNAs in cell differentiation and patterning. Semin Cell Dev Biol. 2008;19:586–595. - PubMed
-
- Shukla LI, Chinnusamy V, Sunkar R. The role of microRNAs and other endogenous small RNAs in plant stress responses. Biochim Biophys Acta. 2008;1779:743–748. - PubMed
-
- Liu PP, et al. Repression of AUXIN RESPONSE FACTOR10 by microRNA160 is critical for seed germination and post-germination stages. Plant J. 2007;52:133–146. - PubMed
-
- Cartolano M, et al. A conserved microRNA module exerts homeotic control over Petunia hybrida and Antirrhinum majus floral organ identity. Nat Genet. 2007;39:901–905. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
