

A designer ligand specific for Kv1.3 channels from a scorpion neurotoxin-based library
- PMID: 20007782
- PMCID: PMC2799710
- DOI: 10.1073/pnas.0910123106
A designer ligand specific for Kv1.3 channels from a scorpion neurotoxin-based library
Abstract
Venomous animals immobilize prey using protein toxins that act on ion channels and other targets of biological importance. Broad use of toxins for biomedical research, diagnosis, and therapy has been limited by inadequate target discrimination, for example, among ion channel subtypes. Here, a synthetic toxin is produced by a new strategy to be specific for human Kv1.3 channels, critical regulators of immune T cells. A phage display library of 11,200 de novo proteins is designed using the alpha-KTx scaffold of 31 scorpion toxin sequences known or predicted to bind to potassium channels. Mokatoxin-1 (moka1) is isolated by affinity selection on purified target. Moka1 blocks Kv1.3 at nanomolar levels that do not inhibit Kv1.1, Kv1.2, or KCa1.1. As a result, moka1 suppresses CD3/28-induced cytokine secretion by T cells without cross-reactive gastrointestinal hyperactivity. The 3D structure of moka1 rationalizes its specificity and validates the engineering approach, revealing a unique interaction surface supported on an alpha-KTx scaffold. This scaffold-based/target-biased strategy overcomes many obstacles to production of selective toxins.
Conflict of interest statement
Conflict of interest statement: Based in part on this work, a patent application (“Identification of Toxin Ligands,” PCT/US2008/013385) has been filed.
Figures
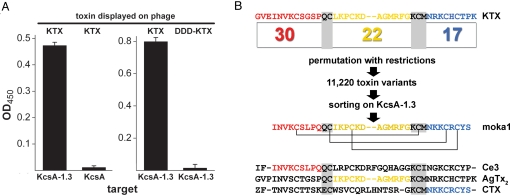
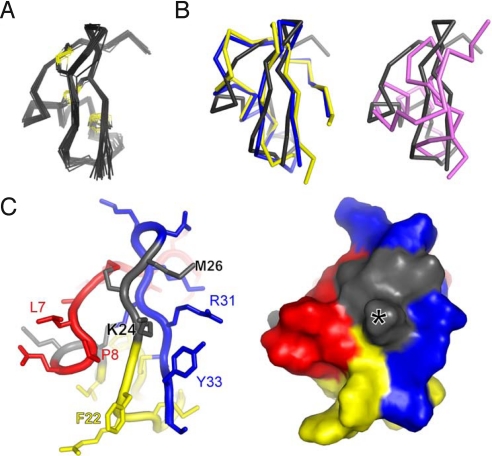
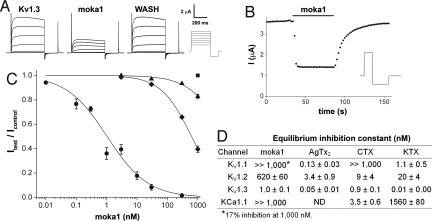
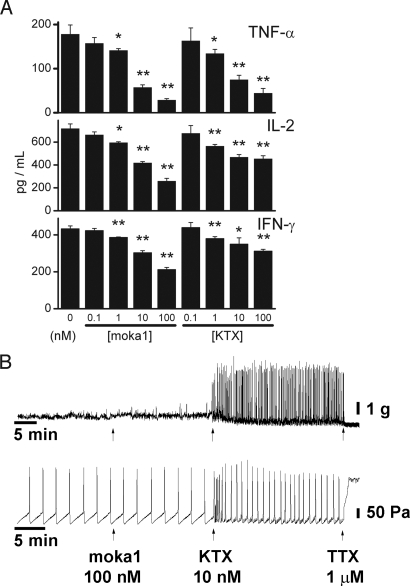
References
-
- Legros C, et al. Engineering-specific pharmacological binding sites for peptidyl inhibitors of potassium channels into KcsA. Biochemistry. 2002;41:15369–15375. - PubMed
-
- Olamendi-Portugal T, et al. Novel α-KTx peptides from the venom of the scorpion Centruroides elegans selectively blockade Kv1.3 over IKCa1 K+ channels of T cells. Toxicon. 2005;46:418–429. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
