

A central role of RLIP76 in regulation of glycemic control
- PMID: 20007934
- PMCID: PMC2828645
- DOI: 10.2337/db09-0911
A central role of RLIP76 in regulation of glycemic control
Abstract
Objective: Pathology associated with oxidative stress frequently results in insulin resistance. Glutathione (GSH) and GSH-linked metabolism is a primary defense against oxidative stress. Electrophilic lipid alkenals, such as 4-hydroxy-t-2-nonenal (4HNE), generated during oxidative stress are metabolized primarily to glutathione electrophile (GS-E) conjugates. Recent studies show that RLIP76 is the primary GS-E conjugate transporter in cells, and a regulator of oxidative-stress response. Because RLIP76(-/-) mice are hypoglycemic, we studied the role of RLIP76 in insulin resistance.
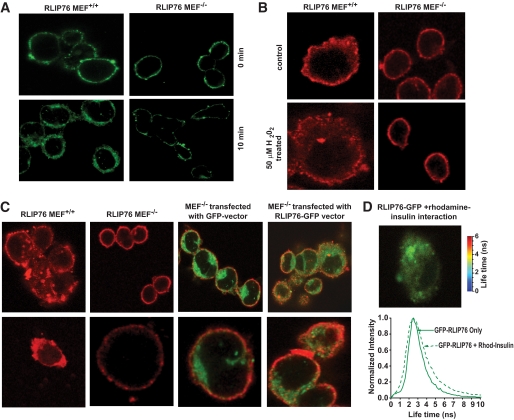
Research design and methods: Blood glucose, insulin, lipid measurements, and hyperinsulinemic-euglycemic and hyperglycemic clamp experiments were performed in RLIP76(+/+) and RLIP76(-/-) C57B mice, using Institutional Animal Care and Use Committee-approved protocols. Time-resolved three-dimensional confocal fluorescence microscopy was used to study insulin endocytosis.
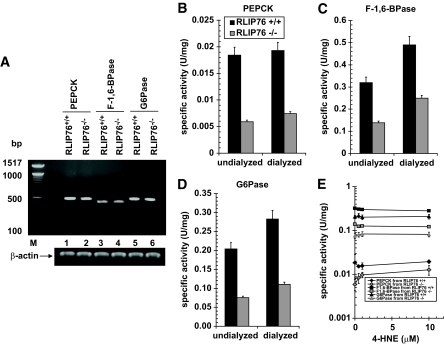
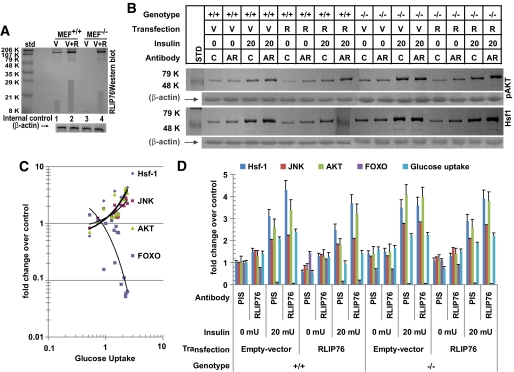
Results: The plasma insulin/glucose ratio was ordered RLIP76(-/-) < RLIP76(+/-) < RLIP76(+/+); administration of purified RLIP76 in proteoliposomes to RLIP76(+/+) animals further increased this ratio. RLIP76 was induced by oxidative or hyperglycemic stress; the concomitant increase in insulin endocytosis was completely abrogated by inhibiting the transport activity of RLIP76. Hydrocortisone could transiently correct hypoglycemia in RLIP76(-/-) animals, despite inhibited activity of key glucocorticoid-regulated hepatic gluconeogenic enzymes, phosphoenolpyruvate carboxykinase, glucose-6-phosphatase, and fructose 1,6-bisphosphatase, in RLIP76(-/-).
Conclusions: The GS-E conjugate transport activity of RLIP76 mediates insulin resistance by enhancing the rate of clathrin-dependent endocytosis of insulin. Because RLIP76 is induced by oxidative stress, it could play a role in insulin resistance seen in pathological conditions characterized by increased oxidative stress.
Figures
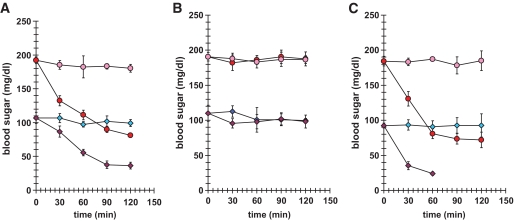
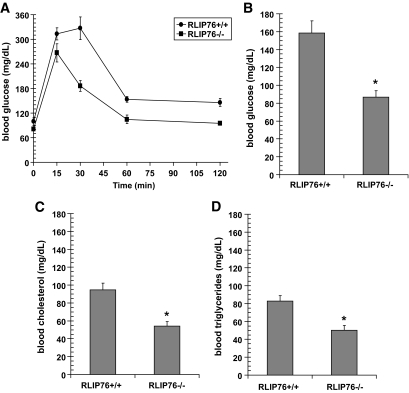
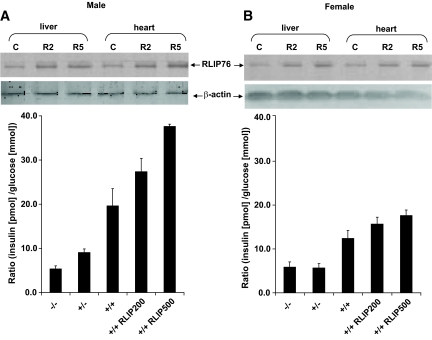
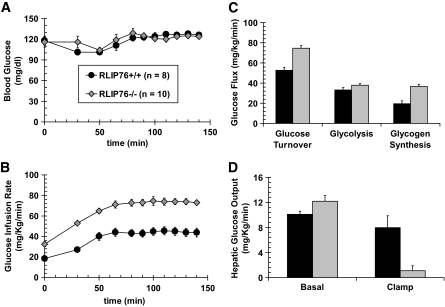
 . Statistical analyses by ANOVA were significant at P < 0.05 for RLIP76+/+ vs. RLIP76−/−; n = 8.
. Statistical analyses by ANOVA were significant at P < 0.05 for RLIP76+/+ vs. RLIP76−/−; n = 8.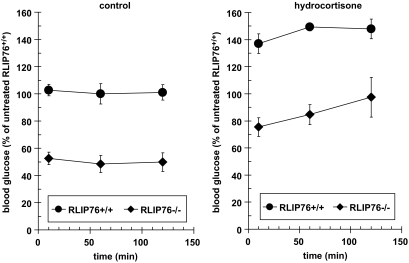

References
-
- Tirosh A, Potashnik R, Bashan N, Rudich A: Oxidative stress disrupts insulin-induced cellular redistribution of insulin receptor substrate-1 and phosphatidylinositol 3-kinase in 3T3–L1 adipocytes: a putative cellular mechanism for impaired protein kinase B activation and GLUT4 translocation. J Biol Chem 1999; 274: 10595– 10602 - PubMed
-
- Awasthi S, Singhal SS, Srivastava SK, Zimniak P, Bajpai KK, Saxena M, Sharma R, Ziller SA, 3rd, Frenkel EP, Singh SV: Adenosine triphosphate-dependent transport of doxorubicin, daunomycin, and vinblastine in human tissues by a mechanism distinct from the P-glycoprotein. J Clin Invest 1994; 93: 958– 965 - PMC - PubMed
-
- Yung LM, Leung FP, Yao X, Chen ZY, Huang Y: Reactive oxygen species in vascular wall. Cardiovasc Hematol Disord Drug Targets 2006; 6: 1– 19 - PubMed
-
- Ceriello A: Acute hyperglycaemia and oxidative stress generation. Diabet Med 1997; 14( Suppl. 3): S45– S49 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
