

Genetic analysis of the regulation of type IV pilus function by the Chp chemosensory system of Pseudomonas aeruginosa
- PMID: 20008072
- PMCID: PMC2812951
- DOI: 10.1128/JB.01390-09
Genetic analysis of the regulation of type IV pilus function by the Chp chemosensory system of Pseudomonas aeruginosa
Abstract
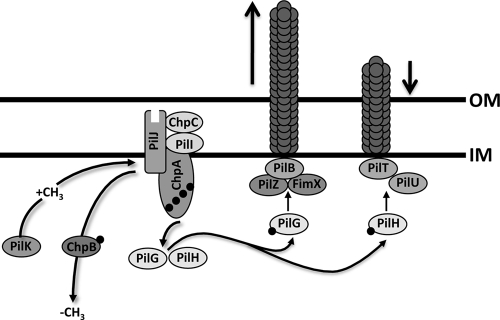
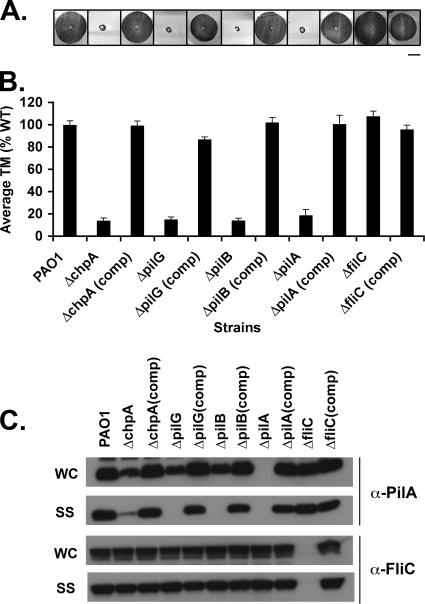
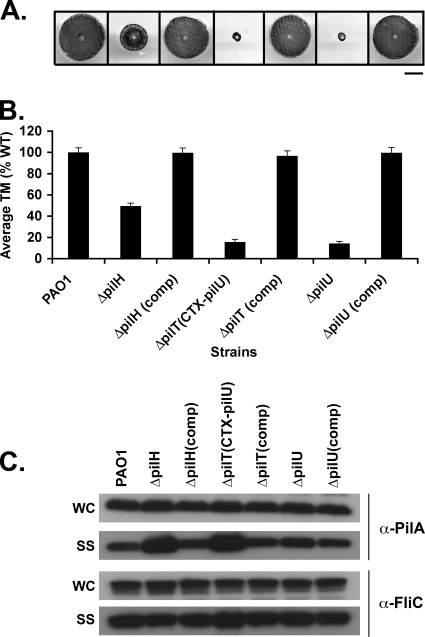
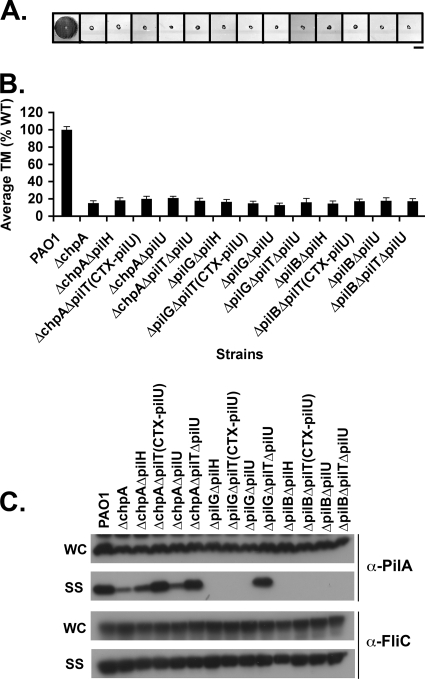
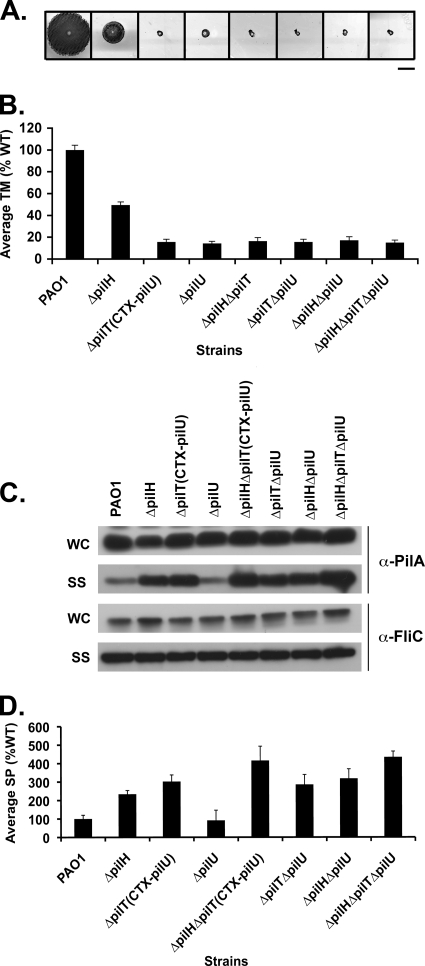
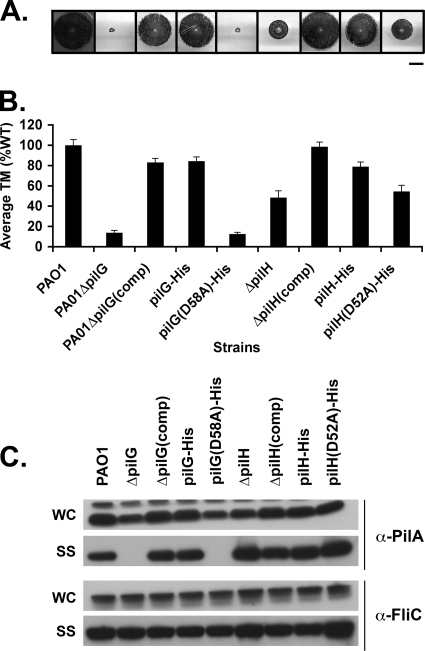
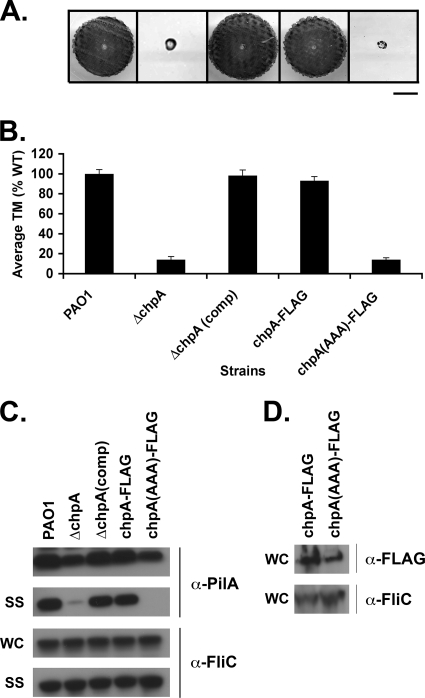
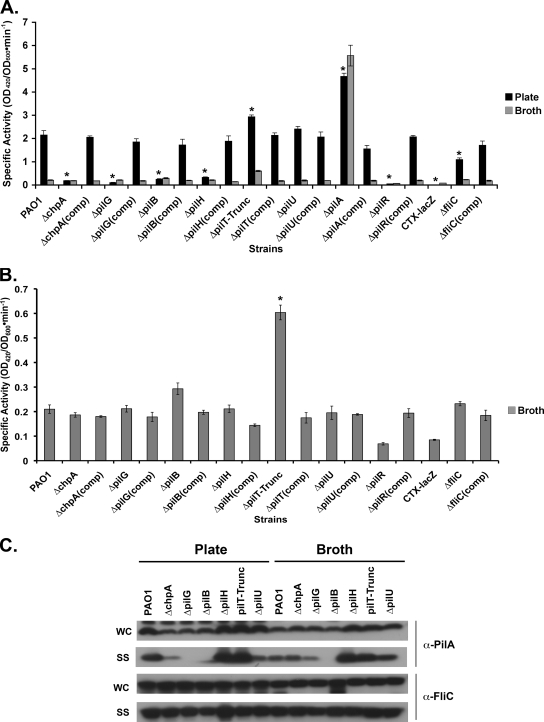
The virulence of the opportunistic pathogen Pseudomonas aeruginosa involves the coordinate expression of many virulence factors, including type IV pili, which are required for colonization of host tissues and for twitching motility. Type IV pilus function is controlled in part by the Chp chemosensory system, which includes a histidine kinase, ChpA, and two CheY-like response regulators, PilG and PilH. How the Chp components interface with the type IV pilus motor proteins PilB, PilT, and PilU is unknown. We present genetic evidence confirming the role of ChpA, PilG, and PilB in the regulation of pilus extension and the role of PilH and PilT in regulating pilus retraction. Using informative double and triple mutants, we show that (i) ChpA, PilG, and PilB function upstream of PilH, PilT, and PilU; (ii) that PilH enhances PilT function; and (iii) that PilT and PilB retain some activity in the absence of signaling input from components of the Chp system. By site-directed mutagenesis, we demonstrate that the histidine kinase domain of ChpA and the phosphoacceptor sites of both PilG and PilH are required for type IV pilus function, suggesting that they form a phosphorelay system important in the regulation of pilus extension and retraction. Finally, we present evidence suggesting that pilA transcription is regulated by intracellular PilA levels. We show that PilA is a negative regulator of pilA transcription in P. aeruginosa and that the Chp system functionally regulates pilA transcription by controlling PilA import and export.
Figures
References
-
- Alm, R., and J. S. Mattick. 1995. Identification of a gene, pilV, required for type 4 fimbrial biogenesis in Pseudomonas aeruginosa, whose product possesses a pre-pilin-like leader sequence. Mol. Microbiol. 16:485-496. - PubMed
-
- Baker, M. D., P. M. Wolanin, and J. B. Stock. 2006. Signal transduction in bacterial chemotaxis. Bioessays 28:9-22. - PubMed
-
- Barken, K. B., S. J. Pamp, L. Yang, M. Gjermansen, J. J. Bertrand, M. Klausen, M. Givskov, C. B. Whitchurch, J. N. Engel, and T. Tolker-Nielsen. 2008. Roles of type IV pili, flagellum-mediated motility and extracellular DNA in the formation of mature multicellular structures in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 10:2331-2343. - PubMed
-
- Becher, A., and H. P. Schweizer. 2000. Integration-proficient Pseudomonas aeruginosa vectors for isolation of single-copy chromosomal lacZ and lux gene fusions. Biotechniques 29:948-952. - PubMed
-
- Bhaya, D., N. R. Bianco, D. Bryant, and A. Grossman. 2000. Type IV pilus biogenesis and motility in the cyanobacterium Synechocystis sp. PCC6803. Mol. Microbiol. 37:941-951. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
